Matemaatiline mudel konservatiivne kiskja saak. Kursusetöö: Kiskja-saagi mudeli kvalitatiivne uurimine. "Predator-Prey" süsteemi simulatsioonimodelleerimine
Kiskjad võivad süüa rohusööjaid ja ka nõrku kiskjaid. Kiskjatel on lai toiduvalik ja nad lülituvad kergesti ühelt saagilt teisele, ligipääsetavamale. Kiskjad ründavad sageli nõrka saaki. Ökoloogiline tasakaal säilib röövloomade-kiskjate populatsioonide vahel.[...]
Kui tasakaal on ebastabiilne (puuduvad piirtsüklid) või väline tsükkel on ebastabiilne, siis mõlema liigi arvukus, mis kogeb tugevat kõikumist, lahkub tasakaalu lähedusest. Veelgi enam, kiire degeneratsioon (esimeses olukorras) toimub röövlooma vähese kohanemise korral, s.t. oma kõrge suremusega (võrreldes ohvri paljunemiskiirusega). See tähendab, et igati nõrk kiskja ei aita kaasa süsteemi stabiliseerumisele ja sureb ise välja.[...]
Eriti tugev on kiskjate surve siis, kui kiskja-saagi koevolutsioonis nihkub tasakaal kiskja poole ja saagi leviala aheneb. Võistlusvõitlus on tihedalt seotud toiduressursside nappusega, see võib olla ka näiteks kiskjate otsene võitlus ruumi kui ressursi pärast, kuid enamasti on see lihtsalt liigi väljatõrjumine, kellel ei ole piisavalt toitu. antud territooriumi poolt liik, kellel on piisavalt samas koguses toitu. See on juba liikidevaheline konkurents.[...]
 |
Ja lõpuks, mudeliga (2.7) kirjeldatud “kiskja-saagi” süsteemis on difusiooniebastabiilsuse tekkimine (kohaliku tasakaalu stabiilsusega) võimalik ainult juhul, kui kiskja loomulik suremus suureneb koos tema populatsiooni kasvuga kiiremini kui lineaarne funktsioon ja troofiline funktsioon erineb Volterra omast või siis, kui saaklooma populatsiooniks on Ollie-tüüpi populatsioon.[...]
Teoreetiliselt võib “üks kiskja - kaks saaklooma” mudelites samaväärne karjatamine (ühe või teise saakloomaliigi eelistamise puudumine) mõjutada röövliikide konkureerivat kooseksisteerimist vaid nendes kohtades, kus potentsiaalselt stabiilne tasakaal juba eksisteerib. Mitmekesisus saab kasvada ainult tingimustes, kus väiksema konkurentsivõimega liikide populatsiooni kasvumäär on suurem kui domineerivatel liikidel. See võimaldab mõista olukorda, kus ühtlane karjatamine toob kaasa taimeliikide mitmekesisuse suurenemise, kus suurem hulk kiireks paljunemiseks valitud liike eksisteerib koos liikidega, mille evolutsioon on suunatud konkurentsivõime tõstmisele.
Samamoodi võib tihedusest sõltuv saakloomade valik viia stabiilse tasakaaluni kahe konkureeriva saakloomaliigi teoreetilistes mudelites, kus tasakaal varem puudus. Selleks peaks kiskja olema võimeline funktsionaalselt ja numbriliselt reageerima saagitiheduse muutustele; võimalik, et vahetus (ebaproportsionaalselt sagedased rünnakud kõige rikkalikuma saagi vastu) on aga olulisem. Tegelikult on kindlaks tehtud, et ümberlülitusel on stabiliseeriv toime süsteemides "üks kiskja - n saak" ja see on ainus mehhanism, mis on võimeline stabiliseerima koostoimeid juhtudel, kui saaklooma nišid kattuvad täielikult. Sellist rolli võivad mängida spetsialiseerimata kiskjad. Spetsialiseerunud röövloomade eelistamine domineerivale konkurendile toimib samamoodi nagu kiskjate vahetamine ja võib stabiliseerida teoreetilisi interaktsioone mudelites, kus varem ei olnud saakliikide vahel tasakaal, eeldusel, et nende nišid on mingil määral eraldatud.[...]
Samuti ei stabiliseeri kiskja “igati tugev” kooslust, s.t. antud saagiks hästi kohanenud ja madala suhtelise suremusega. Sel juhul on süsteemil ebastabiilne piirtsükkel ja hoolimata tasakaaluasendi stabiilsusest degenereerub see juhuslikus keskkonnas (kiskja sööb saagi ära ja selle tulemusena sureb). Selline olukord vastab aeglasele degeneratsioonile.[...]
Seega kiskja hea kohanemise korral stabiilse tasakaalu läheduses võivad tekkida ebastabiilsed ja stabiilsed tsüklid, s.t. Olenevalt algtingimustest kipub “kiskja-saagi” süsteem kas tasakaalu minema või võnkudes eemaldub sellest või tekib tasakaalu läheduses mõlema liigi arvukuse stabiilne kõikumine. [...]
Kiskjatena liigitatud organismid toituvad teistest organismidest, hävitades nende saagi. Seega tuleks elusorganismide hulgas eristada veel ühte klassifikatsioonisüsteemi, nimelt “kiskjad” ja “saakloomad”. Selliste organismide vahelised suhted on kujunenud kogu meie planeedi elu evolutsiooni vältel. Röövorganismid toimivad saakorganismide arvukuse loomulike regulaatoritena. "Riskjate" arvukuse suurenemine toob kaasa "saakloomade" arvukuse vähenemise, mis omakorda vähendab "kiskjate" toiduga varustamist ("saakloomad"), mis üldiselt dikteerib arvukuse vähenemist. “saakloomadest” jne. Seega toimub biotsenoosis röövloomade ja saakloomade arvukuse kõikumised pidevalt, kuid üldiselt tekib teatud tasakaal teatud perioodiks üsna stabiilsete keskkonnatingimuste raames.[... ]
See saavutab lõpuks ökoloogilise tasakaalu röövloomade ja saakloomade populatsioonide vahel.[...]
Kolmandat tüüpi troofilise funktsiooni korral on tasakaaluolek stabiilne, kui kus N on funktsiooni käändepunkt (vt joonis 2, c). See tuleneb sellest, et ajavahemiku jooksul on troofiline funktsioon nõgus ja seetõttu suureneb röövloomade saagitarbimise suhteline osa.[...]
Olgu Гг = -Г, st. on "kiskja-saagi" tüüpi kooslus. Sel juhul on avaldise (7.4) esimene liige võrdne nulliga ja stabiilsuse tingimuse rahuldamiseks tasakaaluseisundi N tõenäosuse osas on nõutav, et ka teine liige ei oleks positiivne.[.. .]
Seega võime vaadeldava kiskja-saaklooma koosluse puhul järeldada, et üldine positiivne tasakaalupositsioon on asümptootiliselt stabiilne, st mis tahes algandmete 1H(0)>0 korral toimub evolutsioon selliselt, et N(7) - ■ K juures tingimusel, et N >0.[...]
Seega hävitab röövloom homogeenses keskkonnas, millel pole sigimiseks peavarju, varem või hiljem saagi populatsiooni ja sureb seejärel ise välja. Elulained” (muutused kiskja ja saaklooma arvukuses) järgnevad üksteisele pideva faasinihkega ning keskmiselt jääb nii kiskja kui ka saaklooma arvukus ligikaudu samale tasemele. Perioodi kestus sõltub mõlema liigi kasvukiirustest ja esialgsetest parameetritest. Saakloomapopulatsiooni jaoks on kiskja mõju positiivne, kuna tema liigne paljunemine tooks kaasa populatsiooni kokkuvarisemise. Kõik mehhanismid, mis takistavad saagi täielikku hävitamist, aitavad omakorda kaasa kiskja toiduvarude säilimisele.[...]
Muud modifikatsioonid võivad olla röövlooma käitumise tagajärg. Saagiks olevate isendite arvul, mida kiskja on võimeline teatud ajahetkel tarbima, on oma piir. Kiskja küllastumise mõju sellele lävele lähenemisel on näidatud tabelis. 2-4, B. Võrranditega 5 ja 6 kirjeldatud vastasmõjudel võivad olla stabiilsed tasakaalupunktid või tsüklilised kõikumised. Sellised tsüklid erinevad aga Lotka-Volterra võrrandites 1 ja 2 kajastatutest. Võrranditega 5 ja 6 esitatud tsüklitel võib olla konstantne amplituud ja keskmine tihedus seni, kuni keskkond on konstantne; pärast häire ilmnemist võivad nad naasta oma varasematele amplituudidele ja keskmisele tihedusele. Selliseid tsükleid, mis häiretest taastuvad, nimetatakse stabiilseteks piirtsükliteks. Jänese ja ilvese vastasmõju võib pidada stabiilseks piirtsükliks, kuid tegemist ei ole Lotka-Volterra tsükliga.[...]
Vaatleme difusiooniebastabiilsuse esinemist kiskja-saagi süsteemis, kuid kõigepealt paneme kirja tingimused, mis tagavad difusiooni ebastabiilsuse esinemise süsteemis (1.1), kus n = 2. On selge, et tasakaal (N, N ) on kohalik (st.[ .. .]
Liigume edasi kiskja ja saaklooma pikaajalise kooseksisteerimisega seotud juhtumite tõlgendamise juurde. On selge, et piirtsüklite puudumisel vastab stabiilne tasakaal populatsiooni kõikumistele juhuslikus keskkonnas ja nende amplituud on võrdeline häirete hajutamisega. See nähtus ilmneb siis, kui röövloomal on kõrge suhteline suremus ja samal ajal kõrge kohanemisvõime antud saagiga.[...]
Vaatleme nüüd, kuidas kiskja sobivuse kasvades muutub süsteemi dünaamika, s.t. b väheneb 1-lt 0-le. Kui sobivus on piisavalt madal, siis piirtsükleid ei ole ja tasakaal on ebastabiilne. Fitnessi suurenemisega selle tasakaalu läheduses võib ilmneda stabiilne tsükkel ja seejärel väline ebastabiilne tsükkel. Olenevalt algtingimustest (kiskja ja saaklooma biomassi suhe) võib süsteem kas kaotada stabiilsuse, s.t. lahkuda tasakaalu lähedusest, vastasel juhul tekivad selles aja jooksul stabiilsed võnked. Fitnessi edasine kasv muudab süsteemi käitumise võnkuva olemuse võimatuks. Kui aga b [...]
Negatiivse (stabiliseeriva) tagasiside näiteks on kiskja ja saagi suhe või ookeani karbonaatsüsteemi toimimine (CO2 lahus vees: CO2 + H2O -> H2CO3). Tavaliselt on ookeanivees lahustunud süsinikdioksiidi kogus osaliselt tasakaalus süsinikdioksiidi kontsentratsiooniga atmosfääris. Süsinikdioksiidi lokaalne suurenemine atmosfääris pärast vulkaanipurskeid põhjustab fotosünteesi intensiivistumist ja selle neeldumist ookeani karbonaadisüsteemis. Kui süsinikdioksiidi tase atmosfääris väheneb, eraldub ookeani karbonaadisüsteem atmosfääri CO2. Seetõttu on süsinikdioksiidi kontsentratsioon atmosfääris üsna stabiilne.[...]
[ ...]
Nagu märkis R. Ricklefs (1979), on olemas tegurid, mis aitavad kaasa suhete stabiliseerimisele "kiskja-saagi" süsteemis: kiskja ebaefektiivsus, alternatiivsete toiduressursside olemasolu kiskja jaoks, vähendades viivitust kiskja ja saagi vahel. kiskja reaktsioon, samuti väliskeskkonna poolt ühele või teisele populatsioonile seatud keskkonnapiirangud. Kiskjate ja saakloomade populatsioonide vastasmõju on väga mitmekesine ja keeruline. Seega, kui kiskjad on piisavalt tõhusad, suudavad nad saaklooma asustustihedust reguleerida, hoides selle alla keskkonna kandevõime. Mõju kaudu, mis neil on röövloomade populatsioonidele, mõjutavad röövloomad saagi erinevate omaduste arengut, mis lõpuks viib ökoloogilise tasakaaluni kiskja ja saaklooma populatsioonide vahel.[...]
Kui üks tingimus on täidetud: 0 1/2. Kui 6 > 1 (kA [...]
Elustiku ja keskkonna stabiilsus sõltub ainult taimede – autotroofide ja taimtoiduliste heterotroofsete organismide koosmõjust. Igas suuruses kiskjad ei ole võimelised rikkuma koosluse ökoloogilist tasakaalu, kuna looduslikes tingimustes ei saa nad oma arvukust konstantse saagiarvuga suurendada. Kiskjad ei pea mitte ainult ise liikuvad olema, vaid saavad toituda ka ainult liikuvatest loomadest.[...]
Ükski teine kala pole nii laialt levinud kui haug. Mõnel üksikul püügipiirkonnal seisvates või voolavates veehoidlates puudub haugi surve saagi ja kiskja vahelise tasakaalu säilitamiseks.Ainult tänapäevased tehisreservuaarid, kus haugid on teiste kalade sigimise tõttu ebasoovitavad kalad, ei ole sihipäraselt asustatud. neid. Haug on maailmas erakordselt hästi esindatud. Neid püütakse kogu põhjapoolkeral USA-st ja Kanadast Põhja-Ameerikas, läbi Euroopa kuni Põhja-Aasiani.[...]
Siin tekib veel üks jätkusuutliku kooseksisteerimise võimalus, suhteliselt kõrge kohanemise kitsas vahemikus. Väga “hea” kiskjaga ebastabiilsele režiimile üleminekul võib tekkida stabiilne väline piirtsükkel, milles biomassi hajumist tasakaalustab selle sissevool süsteemi (saagi kõrge tootlikkus). Siis tekib kurioosne olukord, kui kõige tõenäolisemad on juhuslike võnkumiste amplituudi kaks iseloomulikku väärtust. Mõned esinevad tasakaalu lähedal, teised - piirtsükli lähedal ja nende režiimide vahel on võimalik rohkem või vähem sagedasi üleminekuid.[...]
Hüpoteetilised populatsioonid, mis käituvad vastavalt joonisel fig. 10,11 A, on näidatud joonisel fig. 10.11,-B, kasutades graafikut, mis näitab röövloomade ja saakloomade populatsioonide suhte dünaamikat ning joonisel fig. 10.11.5 kiskjate ja saakloomade arvukuse dünaamika graafikuna ajas. Saakloomapopulatsioonis, kui see liigub madala tihedusega tasakaalust suure tihedusega tasakaalu ja naaseb tagasi, toimub arvude "puhang". Ja see arvude plahvatuslik suurenemine ei ole keskkonna samaväärse muutuse tagajärg. Vastupidi, see arvumuutus tuleneb mõjust endast (väikese "müraga" keskkonnas) ja eelkõige peegeldab see mitme tasakaaluseisundi olemasolu. Sarnaseid arutluskäike saab kasutada ka keerukamate populatsioonidünaamika juhtumite selgitamiseks looduslikes populatsioonides.[...]
Ökosüsteemi kõige olulisem omadus on selle stabiilsus, vahetuse tasakaal ja selles toimuvad protsessid. Populatsioonide või ökosüsteemide võimet säilitada muutuvates keskkonnatingimustes stabiilset dünaamilist tasakaalu nimetatakse homöostaasiks (homoios – sama, sarnane; staas – olek). Homöostaas põhineb tagasiside põhimõttel. Looduses tasakaalu säilitamiseks pole välist kontrolli vaja. Homöostaasi näiteks on alamsüsteem “kiskja-saakloom”, milles reguleeritakse kiskja ja saaklooma asustustihedust.[...]
Looduslik ökosüsteem (biogeocenoos) toimib stabiilselt selle elementide pideva vastasmõju, ainete ringluse, keemilise, energeetilise, geneetilise ja muu energia ning teabe edasikandumise kaudu kettide-kanalite kaudu. Tasakaaluprintsiibi kohaselt kipub iga looduslik süsteem, mille energia- ja infovoog seda läbib, arenema püsiseisundisse. Samas tagatakse tagasisidemehhanismi kaudu automaatselt ökosüsteemide stabiilsus. Tagasiside seisneb ökosüsteemide hallatavatelt komponentidelt saadud andmete kasutamises, et halduskomponentide poolt protsessi kohandada. Eespool selles kontekstis käsitletud "kiskja"-"saaklooma" suhet saab kirjeldada mõnevõrra üksikasjalikumalt; Seega söövad veeökosüsteemis röövkalad (tiigis haug) teisi röövkalaliike (ristkarpkala); kui ristikarpkala arvukus suureneb, on see näide positiivsest tagasisidest; haug, kes toitub ristikarpkaladest, vähendab selle arvukust - see on näide negatiivsest tagasisidest; kiskjate arvukuse kasvades langeb ohvrite arv ning toidupuuduses olev kiskja vähendab ka oma populatsiooni kasvu; lõpuks saavutatakse kõnealuses tiigis dünaamiline tasakaal nii haugi kui ka karpkala arvukuses. Pidevalt hoitakse tasakaalu, mis välistaks troofilise ahela mistahes lüli kadumise (joonis 64).[...]
Liigume edasi kõige olulisema üldistuse juurde, nimelt, et negatiivsed vastasmõjud muutuvad aja jooksul vähem märgatavaks, kui ökosüsteem on piisavalt stabiilne ja selle ruumiline struktuur võimaldab populatsioonide vastastikust kohanemist. Lotka-Volterra võrrandiga kirjeldatud kiskja-saaklooma tüüpi mudelsüsteemides, kui võrrandisse ei sisestata arvude isepiiravate tegurite toimet iseloomustavaid lisatermineid, tekivad võnked pidevalt ega sure välja (vt Lewontin , 1969). Pimentel (1968; vt ka Pimentel ja Stone 1968) näitas eksperimentaalselt, et sellised lisaterminid võivad peegeldada vastastikust kohanemist või geneetilist tagasisidet. Kui indiviididest loodi uusi kultuure, kes olid varem kaks aastat koos eksisteerinud kultuuris, kus nende arvukus kõikus, ilmnes, et neil tekkis ökoloogiline homöostaas, kus iga populatsioon "suruti" teise poolt selliseks. niivõrd, et osutus võimalikuks nende kooseksisteerimine stabiilsemas tasakaalus.
Kolmogorovi mudel teeb ühe olulise eelduse: kuna eeldatakse, et see tähendab, et saakloomapopulatsioonis on mehhanismid, mis reguleerivad nende arvukust ka kiskjate puudumisel.
Kahjuks ei võimalda selline mudeli sõnastus meil vastata küsimusele, mille üle on viimasel ajal palju vaieldud ja mida me juba peatüki alguses mainisime: kuidas saab röövloomade populatsioon avaldada regulatiivset mõju piirkonna populatsioonile. saagiks, et kogu süsteem oleks jätkusuutlik? Seetõttu pöördume tagasi mudeli (2.1) juurde, milles saakloomapopulatsioonis (nagu ka kiskjate populatsioonis) puuduvad eneseregulatsiooni mehhanismid (näiteks regulatsioon liigisisese konkurentsi kaudu); seetõttu on kooslusse kuuluvate liikide arvukuse reguleerimise ainsaks mehhanismiks kiskjate ja saaklooma vaheline troofiline suhe.
Siin (nii et erinevalt eelmisest mudelist sõltuvad lahendused (2.1) loomulikult konkreetsest troofilise funktsiooni tüübist, mille omakorda määrab röövloomade olemus, st kiskja troofiline strateegia ja kaitsestrateegia Kõigile nendele funktsioonidele (vt joonis I) on ühised järgmised omadused:
Süsteemil (2.1) on üks mittetriviaalne statsionaarne punkt, mille koordinaadid määratakse võrranditest
![]()
loomuliku piirangu all.
On veel üks statsionaarne punkt (0, 0), mis vastab triviaalsele tasakaalule. Lihtne on näidata, et see punkt on sadul ja eraldajad on koordinaatteljed.
Punkti iseloomulikul võrrandil on vorm
![]()
Ilmselgelt klassikalise Volterra mudeli jaoks.
Seetõttu võib f väärtust pidada vaadeldava mudeli Volterra mudelist kõrvalekaldumise mõõdupuuks.
![]()
fookus on statsionaarne punkt ja süsteemis ilmnevad võnked; kui vastupidine ebavõrdsus on täidetud, on sõlm olemas ja süsteemis pole võnkeid. Selle tasakaaluseisundi stabiilsuse määrab tingimus
st see sõltub oluliselt kiskja troofilise funktsiooni tüübist.
Tingimust (5.5) saab tõlgendada järgmiselt: kiskja-saaklooma süsteemi mittetriviaalse tasakaalu stabiilsuseks (ja seega ka selle süsteemi olemasoluks) piisab sellest, et selle oleku läheduses on tarbitud saagi suhteline osakaal. kiskja poolt saakloomade arvu suurenedes. Tõepoolest, röövlooma tarbitud saaklooma osa (koguarvust) kirjeldab diferentseeruv funktsioon, mille suurendamise tingimus (positiivne tuletis) näeb välja selline
![]()
Viimane punktis võetud tingimus pole midagi muud kui tasakaalu stabiilsuse tingimus (5.5). Järjepidevuse juures peab see täituma ka punkti teatud naabruses Seega, kui ohvrite arv selles naabruses, siis
Olgu nüüd troofilisel funktsioonil V joonisel fig. 11, a (selgrootutele iseloomulik). Võib näidata, et kõigi lõplike väärtuste korral (kuna see on ülespoole kumer)
see tähendab, et ohvrite statsionaarse arvu mis tahes väärtuse korral ei ole ebavõrdsus (5,5) täidetud.
See tähendab, et seda tüüpi troofilise funktsiooniga süsteemis ei ole stabiilset mittetriviaalset tasakaalu. Võimalikud on mitmesugused tagajärjed: kas nii saakloomade kui ka kiskjate arvukus suureneb lõputult või (kui trajektoor möödub ühe koordinaattelje lähedalt) juhuslikel põhjustel muutub saaklooma või kiskjate arv nulliks. Kui saakloom sureb, sureb mõne aja pärast ka kiskja, kui aga kiskja sureb esimesena, hakkab saagi arvukus plahvatuslikult kasvama. Kolmas võimalus - stabiilse piirtsükli tekkimine - on võimatu, mida on lihtne tõestada.
Tegelikult väljend

positiivses kvadrandis on alati positiivne, välja arvatud juhul, kui sellel on joonisel fig. 11, a. Siis ei ole Dulaci kriteeriumi järgi selles piirkonnas suletud trajektoore ja stabiilset piirtsüklit ei saa eksisteerida.
Seega võime järeldada: kui troofilisel funktsioonil on joonisel fig. 11, ja siis ei saa kiskja olla regulaator, mis tagab saaklooma populatsiooni stabiilsuse ja seeläbi kogu süsteemi kui terviku stabiilsuse. Süsteem saab olla stabiilne ainult siis, kui saaklooma populatsioonil on oma sisemised regulatsioonimehhanismid, näiteks liigisisene konkurents või episootia. Seda reguleerimise võimalust on juba käsitletud §-des 3, 4.
Varem märgiti, et seda tüüpi troofiline funktsioon on iseloomulik röövputukatele, kelle "ohvriteks" on tavaliselt ka putukad. Teisest küljest näitavad paljude „kiskja-saaklooma” tüüpi looduslike koosluste, sealhulgas putukaliikide dünaamika vaatlused, et neid iseloomustavad väga suure amplituudiga ja väga spetsiifilist tüüpi kõikumised.
Tavaliselt toimub pärast enam-vähem järkjärgulist arvude suurenemist (mis võib toimuda kas monotoonselt või võnkumiste kujul koos kasvava amplituudiga) järsk langus (joon. 14) ja siis pilt kordub. Ilmselt võib putukaliikide arvukuse dünaamika olemust seletada selle süsteemi ebastabiilsusega väikese ja keskmise arvukuse korral ning võimsate populatsioonisisese arvukuse regulaatorite toimega suurel arvul.

Riis. 14. Eukalüptipuudest toituva Austraalia psülliidi Cardiaspina albitextura populatsiooni dünaamika. (Artiklist: Clark L. R. Cardiaspina albitextura populatsioonidünaamika.-Austr. J. Zool., 1964, 12, nr 3, lk 362-380.)
Kui “kiskja-saagi” süsteemi kuuluvad liigid, mis on võimelised käituma üsna keeruliselt (näiteks kiskjad on õppimisvõimelised või saakloomad on võimelised varjupaika leidma), siis võib sellises süsteemis eksisteerida stabiilne mittetriviaalne tasakaal. See väide on üsna lihtsalt tõestatud.
Tegelikult peaks troofilise funktsiooni vorm olema joonisel fig. 11, c. Selle graafiku punkt on troofilise funktsiooni graafiku alguspunktist tõmmatud sirge puutepunkt.Ilmselt selles punktis on funktsioonil maksimum. Samuti on lihtne näidata, et tingimus (5.5) on kõigi jaoks täidetud. Järelikult on mittetriviaalne tasakaal, kus ohvrite arv on väiksem, asümptootiliselt stabiilne
Siiski ei saa me midagi öelda selle kohta, kui suur on selle tasakaalu stabiilsuspiirkond. Näiteks kui on ebastabiilne piirtsükkel, peab see piirkond asuma tsükli sees. Või teine variant: mittetriviaalne tasakaal (5.2) on ebastabiilne, kuid on olemas stabiilne piirtsükkel; sel juhul saame rääkida ka kiskja-saaklooma süsteemi stabiilsusest. Kuna avaldis (5.7) valides troofilise funktsiooni nagu joonis fig. 11, in saab muuta märki, kui vahetatakse kell , siis Dulaci kriteerium siin ei tööta ja küsimus piirtsüklite olemasolust jääb lahtiseks.

Isendite vastastikmõju kiskja-saakloomade süsteemis
5. kursuse üliõpilane 51 A rühm
Bioökoloogia osakond
Nazarova A. A.
Teadusnõustaja:
Podšivalov A.A.
Orenburg 2011
SISSEJUHATUS
SISSEJUHATUS
Igapäevastes arutlustes ja tähelepanekutes juhindume me ise seda teadmata ja sageli isegi aru saamata aastakümneid tagasi avastatud seadustest ja ideedest. Arvestades kiskja-saagi probleemi, oletame, et ohver mõjutab kaudselt ka kiskjat. Mis oleks lõvil õhtusöögiks, kui antiloope poleks; mida teeksid juhid, kui töötajaid poleks; kuidas arendada äri, kui klientidel raha pole...
"Kiskja-saakloom" süsteem on keeruline ökosüsteem, mille puhul on realiseeritud pikaajalised suhted röövloomade ja saakloomade vahel, mis on tüüpiline koevolutsiooni näide. Suhe kiskjate ja nende saagi vahel areneb tsükliliselt, näitlikustades neutraalset tasakaalu.
Selle liikidevaheliste suhete vormi uurimine võimaldab lisaks huvitavate teaduslike tulemuste saamisele lahendada palju praktilisi probleeme:
biotehniliste meetmete optimeerimine nii saakliikide kui ka kiskjate suhtes;
territoriaalkaitse kvaliteedi parandamine;
küttimissurve reguleerimine jahimaadel jne.
Eeltoodu määrab valitud teema asjakohasuse.
Kursusetöö eesmärk on uurida isendite interaktsiooni “kiskja-saaklooma” süsteemis. Eesmärgi saavutamiseks püstitatakse järgmised ülesanded:
kisklus ja selle roll troofiliste suhete kujunemisel;
kiskja-saagi suhte põhimudelid;
sotsiaalse elustiili mõju "kiskja-saagi" süsteemi stabiilsusele;
kiskja-saakloomade süsteemi laboratoorne modelleerimine.
Kiskjate mõju saakloomade arvukusele ja vastupidi on üsna ilmne, kuid selle koostoime mehhanismi ja olemuse kindlaksmääramine on üsna keeruline. Kavatsen neid probleeme oma kursusetöös käsitleda.
#�������####################################### ### #######"#5#@#?#8#;#0###��####################+# #### ######��\############### ###############��#���# #### ######## 4. peatükkPEATÜKK 4. SÜSTEEMI “PREDATOR – PRIMIT” LABORATOORNE MODELLEERIMINE
Duke'i ülikooli teadlased on koostöös kolleegidega Stanfordi ülikoolist, Howard Hughesi meditsiiniinstituudist ja California tehnoloogiainstituudist dr Lingchong You juhtimisel välja töötanud geneetiliselt muundatud bakterite elava süsteemi, mis võimaldab üksikasjalikumalt uurida. kiskja-saakloomade vastasmõju populatsiooni tasandil.
Uus eksperimentaalne mudel on näide kunstlikust ökosüsteemist, milles teadlased programmeerivad baktereid uusi funktsioone täitma. Sellised ümberprogrammeeritud bakterid leiavad laialdast rakendust meditsiinis, keskkonna puhastamisel ja bioarvutite loomisel. Selle töö raames kirjutasid teadlased E. coli (Escherichia coli) “tarkvara” ümber nii, et kaks erinevat bakteripopulatsiooni moodustasid laboris tüüpilise kiskja-saagi interaktsiooni süsteemi, mille eripäraks oli see, et bakterid ei ahminud üksteist, vaid kontrollisid vastase populatsiooni, muutes “enesetappude” sagedust.
Sünteetilise bioloogiana tuntud uurimisvaldkond tekkis umbes 2000. aastal ja enamik sellest ajast alates loodud süsteeme tugineb ühe bakteri ümberprogrammeerimisele. Autorite väljatöötatud mudel on ainulaadne selle poolest, et see koosneb kahest samas ökosüsteemis elavast bakteripopulatsioonist, mille ellujäämine sõltub üksteisest.
Sellise süsteemi eduka toimimise võti on kahe populatsiooni võime üksteisega suhelda. Autorid lõid kaks bakteritüve – “kiskjad” ja “taimsööjad”, mis olenevalt olukorrast eraldavad üldisesse ökosüsteemi toksilisi või kaitsvaid ühendeid.
Süsteemi tööpõhimõte põhineb röövloomade arvukuse ja saaklooma suhte säilitamisel kontrollitud keskkonnas. Rakkude arvu muutused ühes populatsioonis aktiveerivad ümberprogrammeeritud geene, mis käivitab teatud keemiliste ühendite sünteesi.
Seega põhjustab väike saakloomade hulk keskkonnas kiskja rakkudes enesehävitusgeeni aktiveerumist ja nende surma. Ohvrite arvu kasvades saavutab nende keskkonda eralduv ühend aga kriitilise kontsentratsiooni ja aktiveerib kiskjageeni, mis annab enesetapugeenile “antidoodi” sünteesi. See toob kaasa kiskjate arvukuse suurenemise, mis omakorda toob kaasa kiskjate sünteesitud ühendi akumuleerumise keskkonda, mis sunnib ohvreid enesetapule.
Fluorestsentsmikroskoopiat kasutades dokumenteerisid teadlased röövloomade ja saaklooma vastastikmõju.
Rohelist värvi kiskjarakud kutsuvad esile punaseks värvitud röövrakkude enesetapu. Saakloomaraku pikenemine ja purunemine viitab selle surmale.
See süsteem ei kujuta täpselt ette kiskja ja saakloomade vastasmõju looduses, sest kiskjabakterid ei toitu saakbakteritest ja mõlemad populatsioonid konkureerivad samade toiduvarude pärast. Siiski usuvad autorid, et nende väljatöötatud süsteem on kasulik vahend bioloogiliste uuringute jaoks.
Uus süsteem näitab selget seost geneetika ja populatsioonidünaamika vahel, mis aitab tulevastes uuringutes molekulaarsete interaktsioonide mõju kohta rahvastiku muutustele, mis on ökoloogia keskne teema. Süsteem pakub praktiliselt piiramatuid muutuvaid manipuleerimisvõimalusi, et uurida üksikasjalikult keskkonna, geeniregulatsiooni ja populatsiooni dünaamika vahelisi koostoimeid.
Seega on bakterite geneetilist aparaati kontrollides võimalik imiteerida keerukamate organismide arengu- ja koostoimeprotsesse.
3. PEATÜKK
3. PEATÜKK. SOTSIAALSE ELUKORDA MÕJU „KISKJA-OHVRI” SÜSTEEMI STABIILSUSELE
USA ja Kanada ökoloogid on näidanud, et kiskjate ja nende saakloomade grupiviisiline eluviis muudab radikaalselt kiskja-saagi süsteemi käitumist ja annab sellele suurema stabiilsuse. See efekt, mida kinnitavad Serengeti pargi lõvide ja gnuude populatsioonidünaamika vaatlused, põhineb lihtsal tõsiasjal, et grupiviisilise eluviisiga väheneb röövloomade ja potentsiaalsete ohvrite juhuslike kohtumiste sagedus.
Ökoloogid on välja töötanud hulga matemaatilisi mudeleid, mis kirjeldavad röövloom-saakloomade süsteemi käitumist. Eelkõige selgitavad need mudelid hästi mõnikord täheldatud järjekindlaid perioodilisi kõikumisi röövloomade ja saakloomade arvukuses.
Selliseid mudeleid iseloomustab tavaliselt kõrge ebastabiilsuse tase. Teisisõnu, nendes mudelites paljude sisendparameetrite (nagu kiskjate suremus, röövloomade biomassi kiskjate biomassiks muundamise efektiivsus jne) korral varem või hiljem kõik kiskjad kas välja või esmalt surevad. süüa kõik ohvrid ära ja siis ikkagi nälga surra.
Looduslikes ökosüsteemides on muidugi kõik keerulisem kui matemaatilises mudelis. Ilmselt on palju tegureid, mis võivad kiskja-saakloomade süsteemi stabiilsust tõsta ja tegelikkuses toob see harva kaasa nii järske arvukuse hüppeid kui Kanada ilvestel ja jänestel.
Kanada ja USA ökoloogid avaldasid ajakirja viimases numbris " loodus" artikkel, mis juhtis tähelepanu ühele lihtsale ja ilmselgele tegurile, mis võib kiskja-saakloomade süsteemi käitumist dramaatiliselt muuta. See puudutab grupielu.
Enamik saadaolevaid mudeleid eeldab röövloomade ja nende saagi ühtlast jaotumist antud piirkonnas. Nende koosolekute sageduse arvutused põhinevad sellel. On selge, et mida suurem on saakloomade tihedus, seda sagedamini kiskjad neid kohtavad. Sellest sõltub rünnakute, sealhulgas edukate rünnakute arv ja lõpuks kiskjate röövloomade intensiivsus. Näiteks kui saaklooma on liiga palju (kui pole vaja otsimisele aega raisata), piirab tarbimise kiirust ainult aeg, mis on vajalik kiskjal järgmise ohvri püüdmiseks, tapmiseks, söömiseks ja seedimiseks. Kui saaki kohtab harva, on peamiseks karjatamise kiirust määravaks teguriks ohvri otsimiseks kuluv aeg.
Kiskja-saakloomade süsteemide kirjeldamiseks kasutatavates ökoloogilistes mudelites mängib võtmerolli karjatamise intensiivsuse (ühe kiskja poolt ajaühikus söödud saakloomade arv) sõltuvuse olemus saaklooma populatsiooni tihedusest. Viimast hinnatakse loomade arvuna pindalaühiku kohta.
Tuleb märkida, et nii saakloomade kui ka kiskjate rühmaelustiili puhul ei täitu esialgne eeldus loomade ühtlasest ruumilisest jaotusest ning seetõttu muutuvad kõik edasised arvutused valeks. Näiteks karja röövloomade elustiili puhul ei sõltu kiskjaga kohtumise tõenäosus tegelikult mitte üksikute loomade arvust ruutkilomeetril, vaid karjade arvust sama pindalaühiku kohta. Kui saak oleks jaotunud ühtlaselt, komistaksid kiskjad neile palju sagedamini kui karja eluviisiga, kuna karjade vahele tekivad suured ruumid, kus saaki pole. Sarnane tulemus saadakse ka röövloomade grupiviisilise eluviisiga. Üle savanni uitav lõvide uhkus ei märka palju rohkem potentsiaalset saaki, kui seda teeks üksildane lõvi, kes kõnnib sama teed.
Kolm aastat (2003–2007) jälgisid teadlased Serengeti pargi (Tansaania) suurel alal lõvisid ja nende saaki (peamiselt gnuud) hoolikalt. Rahvastikutihedust registreeriti iga kuu; Regulaarselt hinnati ka lõvide erinevate sõraliste liikide tarbimise intensiivsust. Nii lõvid ise kui ka seitse peamist saagiliiki juhivad grupiviisilist eluviisi. Selle asjaolu arvessevõtmiseks tegid autorid vajalikud muudatused standardsetesse keskkonnavalemitesse. Mudelid parameetriseeriti vaatluste käigus saadud reaalsete kvantitatiivsete andmete põhjal. Arvesse võeti nelja mudeli varianti: esimeses eirati röövloomade ja saaklooma grupiviisilist eluviisi, teises arvestati seda ainult kiskjate, kolmandas ainult saaklooma ja neljandas mõlema puhul. .
|
|
Nagu arvata võis, vastas tegelikkusele kõige paremini neljas variant. Ta osutus ka kõige stabiilsemaks. See tähendab, et selle mudeli sisendparameetrite laia valikuga on kiskjate ja saaklooma pikaajaline stabiilne kooseksisteerimine võimalik. Pikaajaliste vaatluste andmed näitavad, et selles osas peegeldab mudel adekvaatselt ka tegelikkust. Lõvide ja nende saakloomade arvukus Serengeti pargis on üsna stabiilne, perioodilisi koordineeritud kõikumisi (nagu ilveste ja jäneste puhul) ei ole täheldatud.
Tulemused näitavad, et kui lõvid ja gnuud elaksid üksi, tooks saakloomade arvu suurenemine kaasa röövloomade kiire kiirenemise. Tänu grupiviisilisele eluviisile seda ei juhtu, kiskjate aktiivsus suureneb suhteliselt aeglaselt ja üldine karjatamise tase jääb madalaks. Autorite sõnul, mida toetavad mitmed kaudsed tõendid, ei piira Serengeti pargi ohvrite arvu mitte lõvid, vaid toiduvarud.
Kui kollektivismi kasu ohvritele on üsna ilmne, siis lõvide osas jääb küsimus lahtiseks. See uuring näitas selgelt, et röövlooma grupiviisilisel eluviisil on tõsine puudus – tegelikult saab iga üksik lõvi seetõttu vähem saaki. Ilmselgelt tuleb seda puudust kompenseerida mõne väga olulise eelisega. Traditsiooniliselt arvati, et lõvide seltskondlik elustiil on seotud suurte loomade küttimisega, kellega on isegi lõvil üksi raske hakkama saada. Viimasel ajal on aga paljud eksperdid (sealhulgas arutluse all oleva artikli autorid) hakanud kahtlema selle seletuse õigsuses. Nende arvates vajavad lõvid ühistegevust ainult pühvlite küttimisel ja lõvid eelistavad teist tüüpi saagiga üksi hakkama saada.
Usutavam tundub oletus, et uhkusi on vaja puhtalt sisemiste probleemide reguleerimiseks, mida lõvi elus on palju. Näiteks on nende seas levinud lapsetapmine – teiste inimeste poegade tapmine isaste poolt. Gruppi jäävatel emastel on lihtsam oma lapsi agressorite eest kaitsta. Lisaks on uhkusel palju lihtsam kui üksikul lõvil kaitsta oma jahipiirkonda naaberpriide eest.
Allikas: John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. Rühma moodustumine stabiliseerib kiskja ja saagi dünaamikat // Loodus. 2007. V. 449. Lk 1041–1043.
Simulatsiooni modelleerimine süsteemid "Kiskja-ohver"
Kokkuvõte >> Majanduslik ja matemaatiline modelleerimine... süsteemid « Kiskja-ohver" Teostaja Gizyatullin R.R gr.MP-30 Kontrollinud Lisovets Y.P. MOSCOW 2007 Sissejuhatus Interaktsioon...mudel interaktsiooni kiskjad Ja ohvrid pinnal. Eelduste lihtsustamine. Proovime võrrelda ohvrile Ja kiskja mõned...
Kiskja-Ohver
Abstraktne >> ÖkoloogiaMatemaatilise ökoloogia rakendused on süsteem kiskja-ohver. Selle tsükliline käitumine süsteemid statsionaarses keskkonnas oli... lisades mittelineaarne interaktsiooni vahel kiskja Ja ohver. Saadud mudelil on ...
Ökoloogia abstraktne
Abstraktne >> Ökoloogiategur ohvrid. Sellepärast interaktsiooni « kiskja–ohver" on oma olemuselt perioodiline ja seda kirjeldatakse süsteem Lotka võrrandid... nihe oluliselt väiksem kui sisse süsteem « kiskja–ohver". Sarnased interaktsiooni on täheldatud ka Batesi miimikas. ...
Kahte tüüpi interaktsiooni mudelid
Volterra hüpoteesid. Analoogia keemilise kineetikaga. Volterra interaktsioonimudelid. Interaktsioonitüüpide klassifikatsioon Konkurents. Kiskja-saak. Liikide vastasmõju üldistatud mudelid . Kolmogorovi mudel. MacArthuri kahe putukaliigi vahelise interaktsiooni mudel. Parameetriline ja Bazykini süsteemi faasiportreed.
Moodsa matemaatilise populatsiooniteooria rajajaks peetakse õigusega itaalia matemaatikut Vito Volterrat, kes töötas välja bioloogiliste koosluste matemaatilise teooria, mille aparaadiks on diferentsiaal- ja integro-diferentsiaalvõrrandid.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris, 1931). Järgnevatel aastakümnetel arenes rahvastiku dünaamika peamiselt kooskõlas selles raamatus väljendatud ideedega. Volterra raamatu venekeelne tõlge ilmus 1976. aastal pealkirja all: "Olemisvõitluse matemaatiline teooria" koos järelsõnaga Yu.M. Svirežev, mis vaatleb matemaatilise ökoloogia arengulugu aastatel 1931–1976.
Volterra raamat on kirjutatud nii, nagu kirjutatakse matemaatika raamatuid. Esmalt sõnastab see mõned eeldused matemaatiliste objektide kohta, mida peaks uurima, ja seejärel viib läbi nende objektide omaduste matemaatilise uuringu.
Volterra uuritud süsteemid koosnevad kahest või enamast tüübist. Mõnel juhul võetakse arvesse kasutatud toiduainete pakkumist. Nende tüüpide koostoimet kirjeldavad võrrandid põhinevad järgmistel mõistetel.
Volterra hüpoteesid
1. Toit on saadaval kas piiramatus koguses või on selle tarnimine aja jooksul rangelt reguleeritud.
2. Iga liigi isendid surevad välja nii, et ajaühikus sureb konstantne osa olemasolevatest isenditest.
3. Röövliigid söövad ohvreid ja ajaühikus on söödud ohvrite arv alati võrdeline nende kahe liigi isendite kohtumise tõenäosusega, s.t. kiskjate arvu ja saaklooma arvu korrutis.
4. Kui toitu on piiratud koguses ja mitu liiki, kes on võimelised seda tarbima, siis on liigi poolt tarbitud toidu osakaal ajaühikus võrdeline selle liigi isendite arvuga, võetuna teatud koefitsiendiga, mis sõltub toidust. liigid (liikidevahelise konkurentsi mudelid).
5. Kui liik toitub piiramatus koguses saadaolevast toidust, on liikide arvu suurenemine ajaühikus võrdeline liigi arvuga.
6. Kui liik toitub piiratud koguses saadavast toidust, siis tema paljunemist reguleerib toidutarbimise määr, s.o. ajaühiku kohta on kasv võrdeline söödud toidukogusega.
Analoogia keemilise kineetikaga
Neil hüpoteesidel on tihedad paralleelid keemilise kineetikaga. Populatsioonidünaamika võrrandites, nagu ka keemilise kineetika võrrandites, kasutatakse “kokkupõrke põhimõtet”, kui reaktsioonikiirus on võrdeline reageerivate komponentide kontsentratsioonide korrutisega.
Tõepoolest, Volterra hüpoteeside kohaselt kiirus protsessi Iga liigi väljasuremine on võrdeline liikide arvuga. Keemilises kineetikas vastab see teatud aine lagunemise monomolekulaarsele reaktsioonile ja matemaatilises mudelis võrrandite paremal pool asuvatele negatiivsetele lineaarliikmetele.
Keemilise kineetika kontseptsioonide kohaselt on kahe aine vastastikmõju bimolekulaarse reaktsiooni kiirus võrdeline nende ainete kokkupõrke tõenäosusega, s.o. nende kontsentratsiooni korrutis. Samamoodi on Volterra hüpoteeside kohaselt röövloomade paljunemiskiirus (saakloomade surm) võrdeline kiskja ja saaklooma isendite kohtumise tõenäosusega, s.o. nende arvude korrutis. Mõlemal juhul esinevad bilineaarsed liikmed mudelisüsteemis vastavate võrrandite paremal pool.
Lõpuks vastavad Volterra võrrandite paremal pool olevad lineaarsed positiivsed liikmed, mis vastavad populatsioonide kasvule piiramatutes tingimustes, keemiliste reaktsioonide autokatalüütilistele tingimustele. See võrrandite sarnasus keemilistes ja keskkonnamudelites võimaldab populatsiooni kineetika matemaatilisel modelleerimisel rakendada samu uurimismeetodeid kui keemiliste reaktsioonide süsteemide puhul.
Interaktsioonitüüpide klassifikatsioon
Volterra hüpoteeside kohaselt kahe liigi koosmõju, mille arvukus x 1 ja x 2 saab kirjeldada võrranditega:
(9.1)
Siin on parameetrid a i - liigi enda kasvukiiruse konstandid, c mina- arvude isepiirangute konstandid (liigisisene konkurents), b ij- liikide vastasmõju konstandid, (i, j= 1,2). Nende koefitsientide märgid määravad interaktsiooni tüübi.
Bioloogilises kirjanduses klassifitseeritakse koostoimed tavaliselt kaasatud mehhanismide järgi. Siin on mitmekesisus tohutu: mitmesugused troofilised vastasmõjud, bakterite ja planktoni vetikate vahelised keemilised vastasmõjud, seente vastasmõjud teiste organismidega, taimeorganismide järjestus, mis on seotud eelkõige konkurentsiga päikesevalguse pärast ja muldade evolutsiooniga jne. See klassifikatsioon tundub ulatuslik.
E . Odum, võttes arvesse V. Volterra pakutud mudeleid, pakkus klassifikatsiooni mitte mehhanismide, vaid tulemuste järgi. Selle klassifikatsiooni järgi tuleks suhteid hinnata positiivseks, negatiivseks või neutraalseks sõltuvalt sellest, kas ühe liigi arvukus teise liigi juuresolekul suureneb, väheneb või jääb muutumatuks. Seejärel saab peamised interaktsioonitüübid esitada tabeli kujul.
LIIKIDE KOOSTAKTIIVSUSE LIIGID
|
SÜMBIOOS |
b 12 ,b 21 >0 |
||
|
KOMENSALISM |
b 12 ,>0, b 21 =0 |
||
|
RÖÖK-OHVER |
b 12 ,>0, b 21 <0 |
||
|
AMENSALISM |
b 12 ,=0, b 21 <0 |
||
|
VÕISTLUS |
b 12 , b 21 <0 |
||
|
NEUTRALISM |
b 12 , b 21 =0 |
Viimane veerg näitab süsteemi (9.1) interaktsioonikoefitsientide märke.
Vaatame peamisi interaktsiooni liike
VÕISTLUSE VÕRRANDID:
Nagu nägime 6. loengus, on võistlusvõrrandid järgmised:
 (9.2)
(9.2)
Statsionaarsed süsteemilahendused:
(1).
![]()
Mis tahes süsteemiparameetrite koordinaatide lähtekoht on ebastabiilne sõlm.
(2).
![]() (9.3)
(9.3)
C statsionaarne olek (9.3) on sadul at a 1 >b 12 /Koos 2 ja
stabiilne sõlm juures a 1 12 /s 2 . See tingimus tähendab, et liik sureb välja, kui tema enda kasvukiirus on väiksem kui teatud kriitiline väärtus.
(3).
![]() (9.4)
(9.4)
C statsionaarne lahus (9.4)¾ sadul kl a 2 >b 21 /c 1 ja stabiilne sõlm aadressil a 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Statsionaarne olek (9.5) iseloomustab kahe konkureeriva liigi kooseksisteerimist ja esindab stabiilset sõlme, kui seos on täidetud:
![]()
See tähendab ebavõrdsust:
b 12
b 21
mis võimaldab sõnastada liikide kooseksisteerimise tingimust:
Populatsioonidevahelise interaktsiooni koefitsientide korrutis on väiksem kui populatsiooni vastasmõju koefitsientide korrutis.
Tõepoolest, olgu kahe vaatlusaluse liigi loomulik kasvumäära 1 ,a 2 on samad. Siis on stabiilsuse jaoks vajalik tingimus
c 2 > b 12 ,c 1 >b 21 .
Need ebavõrdsused näitavad, et ühe konkurendi suuruse kasv pärsib tema enda kasvu rohkem kui teise konkurendi kasv. Kui mõlema liigi arvukus on osaliselt või täielikult piiratud erinevate ressurssidega, kehtivad ülaltoodud ebavõrdsused. Kui mõlemal liigil on täpselt samad vajadused, on üks neist elujõulisem ja tõrjub oma konkurendi välja.
Süsteemi faasitrajektooride käitumine annab selge ettekujutuse konkurentsi võimalikest tulemustest. Võrdlustame süsteemi (9.2) võrrandite parempoolsed küljed nulliga:
x 1 (a 1-c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (a 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
Sel juhul saame võrrandid süsteemi peamiste isokliinide jaoks
x 2 = – b 21 x 1 / c 2 +a 2 /c 2, x 2 = 0
– vertikaalsete puutujate isokliinide võrrandid.
x 2 = – c 1 x 1 / b 12+ a 1 /b 12 , x 1 = 0
– vertikaalsete puutujate isokliinide võrrandid. Vertikaalsete ja horisontaalsete puutujasüsteemide isokliinide paarilise lõikepunktid kujutavad võrrandisüsteemi (9.2.) statsionaarseid lahendeid ja nende koordinaate. ![]() on konkureerivate liikide statsionaarne arv.
on konkureerivate liikide statsionaarne arv.
Peamiste isokliinide võimalik asukoht süsteemis (9.2) on näidatud joonisel 9.1. Riis. 9.1Avastab liigi ellujäämiselex 1, joon. 9.1 b– liigi ellujääminex 2, joon. 9.1 V– liikide kooseksisteerimine, kui tingimus (9.6) on täidetud. Joonis 9.1Gdemonstreerib päästikusüsteemi. Siin sõltub võistluse tulemus algtingimustest. Mõlema tüübi nullist erinev statsionaarne olek (9.5) on ebastabiilne. See on sadul, millest separatrix läbib, eraldades iga liigi ellujäämispiirkonnad.

Riis. 9.1.Peamiste isokliinide asukoht kahte tüüpi (9.2) erineva parameetrite suhtega võistlussüsteemi Volterra faasiportreel. Selgitused tekstis.
Liikide konkurentsi uurimiseks viidi läbi katseid väga erinevate organismidega. Tavaliselt valitakse välja kaks lähedalt seotud liiki ning neid kasvatatakse koos ja eraldi rangelt kontrollitud tingimustes. Teatud ajavahemike järel viiakse läbi täielik või valikuline rahvaloendus. Mitmete korduskatsete andmed registreeritakse ja analüüsitakse. Uuringuid viidi läbi algloomadega (eriti ripslastega), paljude Tribolium perekonna mardikaliikidega, drosophila ja magevee koorikloomadega (daphnia). Mikroobipopulatsioonidega on tehtud palju katseid (vt loeng 11). Katseid tehti ka looduses, sealhulgas planaariadega (Reynolds), kahe sipelgaliigiga (Pontin) jne. Joonisel fig. 9.2. kujutab sama ressurssi kasutavate (sama ökoloogilist niši hõivavate) ränivetikate kasvukõveraid. Kui kasvatatakse monokultuuris Asterionella Formosa saavutab konstantse tiheduse taseme ja hoiab ressursi (silikaadi) kontsentratsiooni pidevalt madalal tasemel. B. Kui kasvatatakse monokultuuris Synedrauina käitub sarnaselt ja hoiab silikaatide kontsentratsiooni veelgi madalamal tasemel. B. Kooskasvatamise ajal (kahes eksemplaris) Synedrauina tõrjub välja Asterionella formosa. Ilmselt Synedra

Riis. 9.2.Võistlus ränivetikates. A - kui kasvatatakse monokultuuris Asterionella Formosa saavutab konstantse tiheduse taseme ja hoiab ressursi (silikaadi) kontsentratsiooni pidevalt madalal tasemel. b - kui kasvatatakse monokultuuris Synedrauina käitub sarnaselt ja hoiab silikaatide kontsentratsiooni veelgi madalamal tasemel. V - koos kasvatamisega (kahes eksemplaris) Synedruina tõrjub välja Asterionella formosa. Ilmselt Synedra võidab konkursi tänu oma võimele substraati täielikumalt ära kasutada (vt ka loeng 11).
G. Gause'i katsed konkurentsi uurimisel on laialt tuntud, näidates ühe konkureeriva liigi ellujäämist ja võimaldades tal sõnastada "konkureeriva välistamise seaduse". Seadus ütleb, et ühes ökoloogilises nišis võib eksisteerida ainult üks liik. Joonisel fig. 9.3. Gause katsete tulemused on esitatud kahe sama ökoloogilise niši (joonis 9.3 a, b) ja erinevates ökoloogilistes niššides (joonis 9.3 c) asuvate parametiumi liigi kohta.

Riis. 9.3. A- Kahe liigi populatsiooni kasvukõverad Parametium üheliigilistes kultuurides. Mustad ringid - P Aurelia, valged ringid – P. Caudatum
b- P Aurelia ja P kasvukõverad. Caudatum segakultuuris.
Gause, 1934
Konkurentsimudelil (9.2) on puudusi, eelkõige järeldub, et kahe liigi kooseksisteerimine on võimalik ainult siis, kui nende arvukust piiravad erinevad tegurid, kuid mudel ei näita, kui suured peavad olema erinevused, et tagada pikaajaline kooselu. . Samas on teada, et muutuvas keskkonnas pikaajaliseks kooseksisteerimiseks on vajalik teatud suurusjärguni ulatuv erinevus. Stohhastiliste elementide lisamine mudelisse (näiteks ressursikasutuse funktsiooni sisseviimine) võimaldab meil neid probleeme kvantitatiivselt uurida.
RÖÖVJA+OHVRI süsteem
 (9.7)
(9.7)
Siin, erinevalt (9.2), märgid b 12 Ja b 21 on erinevad. Nagu konkurentsi puhul, päritolu
![]() (9.8)
(9.8)
on ebastabiilse sõlme tüüpi spetsiaalne punkt. Kolm muud võimalikku püsiseisundit:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Seega on võimalik ellu jääda ainult saakloom (9,10), ainult kiskja (9,9) (kui tal on muid toiduallikaid) ja mõlema liigi kooselu (9,11). Viimast võimalust oleme juba käsitlenud 5. loengus. Võimalikud faasiportreed kiskja-saakloomade süsteemi jaoks on toodud joonisel fig. 9.4.
Horisontaalsete puutujate isokliinid on sirged
x 2 = – b 21 X 1 /c 2 + a 1/c 2, X 2 = 0,
ja vertikaalsete puutujate isokliinid- sirge
x 2 = – c 1 X 1 /b 12 + a 2 /b 12 , X 1 = 0.
Statsionaarsed punktid asuvad vertikaalsete ja horisontaalsete puutujate isokliinide ristumiskohas.

Jooniselt fig. 9.4 on näha järgmine. Kiskja-saakloomade süsteem (9.7) võib omada stabiilset tasakaaluasendit, milles o Ohvrite rummipopulatsioon suri täielikult välja ( ) ja alles jäid ainult kiskjad (periood 2 joonisel fig. 9.4 A). Ilmselgelt saab selline olukord realiseeruda ainult siis, kui lisaks kõnealuste ohvrite tüübile X 1 kiskja X 2 – omab täiendavaid toiteallikaid. Seda asjaolu peegeldab mudelis x2 võrrandi paremal pool olev positiivne liige. Erilised punktid(1) ja (3) (joonis 9.4 A) on ebastabiilsed. Teine võimalus – stabiilne statsionaarne seisund, kus kiskjate populatsioon on täielikult välja surnud ja alles on jäänud vaid saak – stabiilne punkt(3) (joonis 9.4 6 ). See on eriline punkt (1) – ka ebastabiilne sõlm.
Lõpuks kolmas võimalus – kiskjate ja saakloomade populatsioonide jätkusuutlik kooseksisteerimine (joonis 1). 9.4 V), mille statsionaarsed arvud on väljendatud valemitega (9.11).
Nagu ühe populatsiooni puhul (vt loeng 3), mudeli puhul (9.7) Stohhastilist mudelit on võimalik välja töötada, kuid seda ei saa otseselt lahendada. Seetõttu piirdume üldiste kaalutlustega. Oletame näiteks, et tasakaalupunkt asub igast teljest teatud kaugusel. Seejärel faasitrajektooride jaoks, millel väärtusedx 1 , x 2 piisavalt suureks, on deterministlik mudel üsna rahuldav. Kuid kui faasitrajektoori mingil hetkel ei ole ükski muutuja väga suur, võivad juhuslikud kõikumised muutuda oluliseks. Need viivad selleni, et esinduspunkt liigub ühele teljele, mis tähendab vastava liigi väljasuremist.
Seega osutub stohhastiline mudel ebastabiilseks, kuna stohhastiline “triiv” viib varem või hiljem ühe liigi väljasuremiseni. Seda tüüpi mudeli puhul sureb kiskja lõpuks välja kas juhuslikult või seetõttu, et tema saagipopulatsioon likvideeritakse kõigepealt. Kiskja-saagi süsteemi stohhastiline mudel selgitab Gause'i katseid hästi (Gause, 1934), milles ripsloomad Paramettum candatum oli ühe teise ripslase ohvriks Didinium nasatum – kiskja. Oodatakse deterministlike võrrandite järgi (9.7) tasakaaluarvud olid neis katsetes igast liigist ligikaudu vaid viis isendit, mistõttu pole üllatav, et igas korduskatses surid kas kiskjad või saakloom (ja pärast neid kiskjad) üsna kiiresti välja. Esitatakse katsete tulemused. joonisel fig. 9.5.

Riis. 9.5. Kõrgus Parametium caudatum ja röövellikud ripsloomad Dadinium nasutum. Alates : Gause G.F. Võitlus olemasolu eest. Baltimore, 1934
Seega näitab Volterra liikide interaktsiooni mudelite analüüs, et vaatamata selliste süsteemide käitumistüüpide mitmekesisusele, ei saa konkureerivate liikide mudelis üldse esineda arvukuses sumbutamata kõikumisi. Selliseid võnkumisi täheldatakse aga looduses ja katsetes. Vajadus nende teoreetilise seletuse järele oli üheks põhjuseks mudelikirjelduste üldisemal kujul formuleerimiseks.
Kahe tüüpi interaktsiooni üldistatud mudelid
Liikide vastasmõju kirjeldamiseks on välja pakutud suur hulk mudeleid, mille võrrandite parempoolsed küljed olid interakteeruvate populatsioonide arvu funktsioonid. Lahendatud sai üldkriteeriumide väljatöötamise küsimus, et määrata kindlaks, millist tüüpi funktsioonid suudavad kirjeldada ajutise populatsiooni suuruse käitumist, sealhulgas stabiilseid kõikumisi. Kõige kuulsamad neist mudelitest kuuluvad Kolmogorovile (1935, muudetud artikkel - 1972) ja Rosenzweigile (1963).
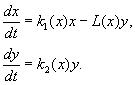 (9.12)
(9.12)
Mudel sisaldab järgmisi eeldusi:
1) Kiskjad ei suhtle omavahel, st. kiskjate paljunemiskiirus k 2 ja ohvrite arv Lühe kiskja poolt ajaühikus hävitatud ei sõltu y.
2) Saaklooma arvu suurenemine kiskjate juuresolekul võrdub kiskjate puudumise suurenemisega, millest on lahutatud röövloomade poolt hävitatud saakloomade arv. Funktsioonid k 1 (x), k 2 (x), L(x), on pidevad ja määratletud positiivsel poolteljel x, y³ 0.
3) dk 1 /dx< 0. See tähendab, et röövloomade paljunemismäär kiskja puudumisel väheneb monotoonselt saakloomade arvukuse suurenemisega, mis peegeldab toidu ja muude ressursside piiratud kättesaadavust.
4) dk 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). Ohvrite arvu suurenemisega väheneb röövloomade paljunemiskoefitsient monotoonselt ohvrite arvu suurenemisega, liikudes negatiivsetelt väärtustelt (kui pole midagi süüa) positiivsetele.
5) Ühe kiskja poolt ajaühikus hävitatud saakloomade arv L(x)> 0 juures N> 0; L(0)=0.
Süsteemi (9.12) faasiportreede võimalikud tüübid on toodud joonisel fig. 9.6:

Riis. 9.6.Kolmogorovi süsteemi faasiportreed (9.12), mis kirjeldab kahe tüübi vastasmõju erinevatel parameetrite vahekordadel. Selgitused tekstis.
Statsionaarsetel lahendustel (neid on kaks või kolm) on järgmised koordinaadid:
(1). ` x=0;` y=0.
Mis tahes parameetri väärtuste koordinaatide alguspunkt on sadul (joonis 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
Amääratakse võrrandist:
k 1 (A)=0.
Statsionaarne lahendus (9.13) on sadul, kui B< A (Joonis 9.6 A, b, G), B võrrandist määratud
k 2 (B)=0
Punkt (9.13) asetatakse positiivsesse kvadranti, kui B>A . See on stabiilne sõlm .
Viimane juhtum, mis vastab kiskja surmale ja saagi ellujäämisele, on näidatud joonisel fig. 9.6 V.
(3). ` x=B,` y=C.(9.14)
C väärtus määratakse võrranditest:
Punkt (9.14) – fookus (joon.9.6 A) või sõlme (joonis 9.6 G), mille stabiilsus sõltub koguse märgists
s 2 = – k 1 (B) – k 1 (B)B+L(B)C.
Kui s>0, punkt on stabiilne, kuis<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
Väliskirjanduses käsitletakse sagedamini Rosenzweigi ja MacArthuri (1963) pakutud sarnast mudelit:
 (9.15)
(9.15)
Kus f(x) - ohvrite arvu muutumise määr x kiskjate puudumisel F( x,y) - röövloomade intensiivsus, k- koefitsient, mis iseloomustab saaklooma biomassi röövloomade biomassiks töötlemise tõhusust, e- kiskjate suremus.
Mudel (9.15) taandub Kolmogorovi mudeli (9.12) erijuhuks järgmistel eeldustel:
1) kiskjate arvukust piirab ainult saakloomade arv,
2) kiirus, millega antud kiskja saaki sööb, sõltub ainult saakloomade populatsiooni tihedusest ja ei sõltu kiskjapopulatsiooni tihedusest.
Seejärel võtavad võrrandid (9.15) kuju.

Reaalsete liikide vastasmõju kirjeldamisel täpsustatakse võrrandite paremad küljed vastavalt arusaamadele bioloogiliste reaalsuste kohta. Vaatleme ühte seda tüüpi populaarseimat mudelit.
Kahe tüüpi putukate vahelise koostoime mudel (MacArthur, 1971)
Mudelit, mida me allpool käsitleme, kasutati kahjulike putukate tõrje praktilise probleemi lahendamiseks, steriliseerides ühe liigi isasloomi. Liikide vastasmõju bioloogiliste tunnuste põhjal kirjutati järgmine mudel
 (9.16)
(9.16)
Siin x,y- kahte tüüpi putukate biomass. Selles mudelis kirjeldatud liikide troofilised vastasmõjud on väga keerulised. See määrab võrrandite paremal küljel olevate polünoomide kuju.
Vaatame esimese võrrandi paremat poolt. Putukate liigid X sööma liigi vastseid juures(liige +k 3 y), kuid liigi täiskasvanud juures sööma liigi vastseid X alluvad suurele liigirikkusele X või juures või mõlemat tüüpi (liikmed -k 4 xy, – y 2). Väikesel X liigi suremus X suurem kui selle loomulik iive (1 -k 1 +k 2 x–x 2 < 0 väikesel X). Teises võrrandis termin k 5 peegeldab liigi loomulikku kasvu y; -k 6 y – seda tüüpi enesepiiramine,-k 7 x– liigi vastsete söömine juures putukate liigid x, k 8 xy – liigi biomassi suurenemine juures liigi täiskasvanud putukate tarbimise tõttu juures liigi vastsed X.
Joonisel fig. 9.7 esitatakse piirtsükkel, mis on süsteemi stabiilse perioodilise lahenduse trajektoor (9.16).

Loomulikult ei saa lahendust küsimusele, kuidas tagada populatsiooni kooseksisteerimine oma bioloogilise keskkonnaga, ilma konkreetse bioloogilise süsteemi eripära ja selle kõigi omavaheliste seoste analüüsita. Samas võimaldab formaalsete matemaatiliste mudelite uurimine vastata mõnele üldisele küsimusele. Võib väita, et selliste mudelite puhul nagu (9.12) ei sõltu populatsioonide kokkusobivuse või mitteühilduvuse fakt nende algsest suurusest, vaid selle määrab ainult liikide interaktsiooni iseloom. Mudel aitab vastata küsimusele: kuidas mõjutada biotsenoosi ja seda juhtida, et kahjulikud liigid kiiresti hävitada.
Juhtimise saab taandada lühiajaliseks järsuks muutuseks populatsiooni väärtustes X Ja u. See meetod vastab tõrjemeetoditele, nagu ühe või mõlema populatsiooni ühekordne hävitamine keemiliste vahenditega. Eespool sõnastatud väitest on selge, et ühilduvate populatsioonide jaoks on see kontrollimeetod ebaefektiivne, kuna aja jooksul jõuab süsteem uuesti statsionaarse režiimi.
Teine võimalus on muuta vaadete interaktsioonifunktsioonide tüüpi, näiteks süsteemiparameetrite väärtuste muutmisel. Just sellele parameetrilisele meetodile vastavad bioloogilised kontrollimeetodid. Seega, kui steriliseeritud isasloomi tuuakse, väheneb loomulik populatsiooni juurdekasv. Kui samal ajal saame teist tüüpi faasiportree, sellise, kus on ainult stabiilne statsionaarne olek nulli kahjurite arvuga, viib kontroll soovitud tulemuseni – kahjuliku liigi populatsiooni hävitamine. Huvitav on märkida, et mõnikord on soovitatav mõju avaldada mitte kahjurile endale, vaid selle partnerile. Üldiselt on võimatu öelda, milline meetod on tõhusam. See sõltub saadaolevatest juhtelementidest ja populatsioonide koostoimet kirjeldavate funktsioonide selgesõnalisest vormist.
A.D. Bazykini mudel
Liikide vastasmõju mudelite teoreetiline analüüs viidi kõige põhjalikumalt läbi A.D. Bazykini raamatus "Interacting Populations Biophysics of Interacting Populations" (M., Nauka, 1985).
Vaatleme üht selles raamatus uuritud kiskja-saagi mudelit.
 (9.17)
(9.17)
Süsteem (9.17) on kõige lihtsama Volterra kiskja-saagi mudeli (5.17) üldistus, mis võtab arvesse kiskja küllastumise mõju. Mudel (5.17) eeldab, et saagikarjatamise intensiivsus suureneb saagitiheduse suurenedes lineaarselt, mis ei vasta tegelikkusele suure saagitiheduse korral. Kiskja toitumise sõltuvuse saagitihedusest kirjeldamiseks saab valida erinevaid funktsioone. Kõige tähtsam on, et valitud funktsioon kasvaks x kaldus asümptootiliselt konstantsele väärtusele. Mudel (9.6) kasutas logistilist sõltuvust. Bazykini mudelis on selliseks funktsiooniks valitud hüperbool x/(1+px). Pidagem meeles, et see on Monodi valemi vorm, mis kirjeldab mikroorganismide kasvukiiruse sõltuvust substraadi kontsentratsioonist. Siin mängib saakloom substraadi rolli ja kiskja mängib mikroorganismide rolli. .
Süsteem (9.17) sõltub seitsmest parameetrist. Parameetrite arvu saab vähendada muutujate asendamisega:
x® (A/D)x; y ® (A/D)/y;
t® (1/A)t; g (9,18)
ja sõltub neljast parameetrist.
Täielikuks kvalitatiivseks uuringuks on vaja neljamõõtmeline parameetriruum jagada erinevat tüüpi dünaamilise käitumisega aladeks, s.t. luua süsteemi parameetriline või struktuurne portree.
Seejärel on vaja konstrueerida faasiportreed iga parameetrilise portree ala jaoks ja kirjeldada faasiportreedega tekkivaid bifurkatsioone parameetrilise portree erinevate alade piiridel.
Täieliku parameetrilise portree konstrueerimine viiakse läbi väikesemõõtmelise parameetrilise portree “lõikude” (projektsioonide) komplektina, millel on mõne parameetri fikseeritud väärtused.
Süsteemi parameetriline portree (9.18) fikseeritud jaoks g ja väike e esitatud joonisel 9.8. Portree sisaldab 10 erinevat tüüpi faasitrajektoori käitumisega ala.

Riis. 9.8.Süsteemi parameetriline portree (9.18) fikseeritud jaoksg
ja väike e
Süsteemi käitumine parameetrite erinevatel suhetel võib olla oluliselt erinev (joonis 9.9). Süsteem võimaldab:
1) üks stabiilne tasakaal (piirkonnad 1 ja 5);
2) üks stabiilne piirtsükkel (piirkonnad 3 ja 8);
3) kaks stabiilset tasakaalu (piirkond 2)
4) stabiilne piirtsükkel ja ebastabiilne tasakaal selle sees (piirkonnad 6, 7, 9, 10)
5) stabiilne piirtsükkel ja stabiilne tasakaal väljaspool seda (piirkond 4).
Parameetrilistes piirkondades 7, 9, 10 piirab tasakaalu tõmbepiirkonda ebastabiilne piirtsükkel, mis asub stabiilses piirkonnas. Kõige huvitavam struktuur on faasiportree, mis vastab parameetrilise portree 6. alale. See on üksikasjalikult näidatud joonisel fig. 9.10.

Tasakaalu B 2 tõmbeala (varjutatud) on ebastabiilsest fookusest B 1 keerduv "tigu". Kui on teada, et algsel ajahetkel asus süsteem B 1 naabruses, siis on võimalik hinnata, kas vastav trajektoor saavutab tasakaalu B 2 või stabiilse piirtsükli, mis ümbritseb kolme tasakaalupunkti C (sadul), B 1 ja B 2 põhinevad tõenäosuslikel kaalutlustel.

Joon.9.10.Süsteemi 9.18 faasiportree parameetrilise piirkonna 6 jaoks. Tõmbepiirkond B 2 on varjutatud
Parameetrilises portrees(9.7) on 22 tekivad mitmesugused bifurkatsioonipiirid 7 erinevat tüüpi bifurkatsioonid. Nende uuring võimaldab meil tuvastada võimalikke süsteemi käitumise tüüpe, kui selle parameetrid muutuvad. Näiteks piirkonnast liikudes 1 alale 3 toimub väikese piirtsükli sünd või enesevõnkumiste pehme sünd ühe tasakaalu ümber IN. Sarnane pehme isevõnkumiste sünd, kuid ümber ühe tasakaalu, nimelt B 1 , tekib piirkondade piiride ületamisel 2 ja 4. Piirkonnast lahkudes 4 alale 5 stabiilne piirtsükkel punkti ümberB 1 "purskab" eraldusribadel ja ainsaks tõmbepunktiks jääb tasakaal B 2 jne.
Praktika jaoks pakub erilist huvi loomulikult süsteemi bifurkatsioonipiiride läheduse kriteeriumide väljatöötamine. Tõepoolest, bioloogid on hästi teadlikud looduslike ökoloogiliste süsteemide "puhverdus" või "paindlikkuse" omadustest. Need terminid viitavad tavaliselt süsteemi võimele absorbeerida välismõjusid. Kuni välismõju intensiivsus ei ületa teatud kriitilist väärtust, ei toimu süsteemi käitumises kvalitatiivseid muutusi. Faasitasandil vastab see süsteemi naasmisele stabiilsesse tasakaaluseisundisse või stabiilsesse piirtsüklisse, mille parameetrid ei erine palju algsest. Kui löögi intensiivsus ületab lubatud piiri, siis süsteem “laguneb” ja läheb kvalitatiivselt erinevale dünaamilise käitumise režiimile, näiteks sureb lihtsalt välja. See nähtus vastab bifurkatsiooni üleminekule.
Igal bifurkatsioonisiirde tüübil on oma eripärad, mis võimaldavad hinnata sellise ülemineku ohtu ökosüsteemile. Siin on mõned üldised kriteeriumid, mis näitavad ohtliku piiri lähedust. Nagu ühe liigi puhul, kui ühe liigi arvukuse vähenemisel süsteem "kinni jääb" ebastabiilse sadulapunkti lähedale, mis väljendub arvu väga aeglases taastamises algväärtusele, siis süsteem on kriitilise piiri lähedal. Ohu indikaatoriks on ka kiskjate ja saakloomade arvukuse kõikumise kuju muutumine. Kui harmoonilisele lähedased võnked muutuvad relaksatsiooniks ja võnkumiste amplituud suureneb, võib see kaasa tuua süsteemi stabiilsuse kaotuse ja ühe liigi väljasuremise.
Liikidevahelise interaktsiooni matemaatilise teooria edasine süvendamine käib populatsioonide endi struktuuri täpsustamise ning ajaliste ja ruumiliste tegurite arvessevõtmise teel.
Kirjandus.
Kolmogorov A.N. Populatsioonidünaamika matemaatiliste mudelite kvalitatiivne uuring. // Küberneetika probleemid. M., 1972, 5. väljaanne.
MacArtur R. Ökoloogiliste süsteemide graafiline analüüs // Division of biology report Perinceton University. 1971
A.D. Bazykin "Interakteeruvate populatsioonide biofüüsika". M., Nauka, 1985.
V. Volterra: "Olelusvõitluse matemaatiline teooria." M.. Teadus, 1976
Gause G.F. Võitlus olemasolu eest. Baltimore, 1934.
Erinevalt (3.2.1) on siin (-012) ja (+a2i) märgid erinevad. Nagu ka konkurentsi puhul (võrrandisüsteem (2.2.1)), on selle süsteemi koordinaatide (1) alguspunkt "ebastabiilse sõlme" tüüpi spetsiaalne punkt. Kolm muud võimalikku püsiseisundit:

Bioloogiline tähendus nõuab positiivseid koguseid X y x 2. Avaldise (3.3.4) puhul tähendab see seda
Kui kiskjate liigisisese konkurentsi koefitsient A,22 = 0, tingimus (3.3.5) viib tingimuseni ai2
Võimalikud faasiportreede tüübid võrrandisüsteemi (3.3.1) jaoks on toodud joonisel fig. 3,2 a-c. Horisontaalsete puutujate isokliinid on sirged
ja vertikaalsete puutujate isokliinid on sirged
Jooniselt fig. 3.2 näitab järgmist. Kiskja-saakloomade süsteemil (3.3.1) võib olla stabiilne tasakaaluasend, kus saakloomade populatsioon on täielikult välja surnud (x = 0) ja alles jäid ainult kiskjad (punkt 2 joonisel 3.26). Ilmselgelt saab selline olukord realiseeruda ainult siis, kui lisaks kõnealuste ohvrite tüübile X kiskja X2 on täiendavaid toiteallikaid. Seda fakti peegeldab mudelis xs võrrandi paremal pool olev positiivne liige. Ainsuse punktid (1) ja (3) (joonis 3.26) on ebastabiilsed. Teine võimalus on stabiilne statsionaarne seisund, kus kiskjate populatsioon on täielikult välja surnud ja alles on jäänud ainult saak - stabiilne punkt (3) (joonis 3.2a). Siin on ainsuse punkt (1) samuti ebastabiilne sõlm.
Kolmandaks võimaluseks on kiskja ja saakloomade populatsioonide stabiilne kooseksisteerimine (joonis 3.2 c), mille statsionaarset arvukust väljendatakse valemitega (3.3.4). Vaatleme seda juhtumit üksikasjalikumalt.
Oletame, et liigisisese konkurentsi koefitsiendid on võrdsed nulliga (ai= 0, i = 1, 2). Oletame ka, et kiskjad toituvad ainult liigi saagist X ja nende puudumisel surevad nad välja kiirusega C2 (punktis (3.3.5) C2).
Uurime seda mudelit üksikasjalikult, kasutades kirjanduses kõige laialdasemalt tunnustatud tähistust. Renoveerimine

Riis. 3.2. Peamiste isokliinide asukoht Volterra süsteemi kiskja-saagi faasiportreel erinevate parameetrite suhete jaoks: A- umbes -
KOOS I C2 C2
1, 3 - ebastabiilne, 2 - stabiilne ainsuse punkt; V -
1, 2, 3 - ebastabiilne, 4 - stabiilne ainsuse punkt, mis on oluline
Kiskja-saagi süsteemil on nendes tähistes järgmine vorm:

Uurime süsteemi (3.3.6) lahenduste omadusi faasitasandil N1
PEAL2
Süsteemil on kaks statsionaarset lahendust. Neid saab hõlpsasti määrata, võrdsustades süsteemi parempoolsed küljed nulliga. Saame: 
Sellest tulenevad statsionaarsed lahendused:

Vaatame teist lahendust lähemalt. Leiame süsteemi (3.3.6) esimese integraali, mis ei sisalda t. Korrutame esimese võrrandi -72-ga, teise -71-ga ja liidame tulemused. Saame:
Nüüd jagame esimese võrrandi arvuga N ja korrutada € 2 ja jagage teine JV-ga 2 ja korrutage arvuga e. Lisame tulemused uuesti:
Võrreldes (3.3.7) ja (3.3.8) saame:

Integreerides saame:

See on soovitud esimene integraal. Seega on süsteem (3.3.6) konservatiivne, kuna sellel on esimene liikumisintegraal, suurus, mis on süsteemi muutujate funktsioon N Ja N2 ja ajast sõltumatu. See omadus võimaldab meil Volterra süsteemide jaoks konstrueerida statistilise mehaanikaga sarnase mõistete süsteemi (vt 5. peatükk), kus süsteemi ajas konstantsel energiaväärtusel on oluline roll.
Iga fikseeritud alates > 0 (mis vastab teatud algandmetele), integraal vastab teatud trajektoorile tasapinnal N1 PEAL2 , mis toimib süsteemi trajektoorina (3.3.6).
Vaatleme Volterra enda pakutud trajektoori koostamise graafilist meetodit. Pange tähele, et valemi (3.3.9) parem pool sõltub ainult D g 2-st ja vasak pool ainult N. Tähistame
(3.3.9) järeldub, et vahel X Ja Y on proportsionaalne suhe
Joonisel fig. 3.3 on näidatud nelja koordinaatsüsteemi esimesed kvadrandid XOY, NOY, N2 HÄRG ja D G 1 0N2 nii et neil kõigil on ühine päritolu.
Ülemises vasakus nurgas (kvadrant EI) funktsiooni graafik (3.3.8) on joonistatud all paremale (kvadrant N2 HÄRG)- funktsioonigraafik Y. Esimesel funktsioonil on min at Ni = ja teine - max at N2 = ?-
Lõpuks kvadrandis XOY konstrueerime sirge (3.3.12) mõne fikseeritud jaoks KOOS.
Märgime punkti N teljel PEAL. See punkt vastab teatud väärtusele Y(N 1), mida on lihtne leida risti joonistades

Riis. 3.3.
läbi N kuni see ristub kõveraga (3.3.10) (vt joonis 3.3). Väärtus K(D^) omakorda vastab kindlale punktile M sirgel Y = cX ja seetõttu mingi väärtus X(N) = JA(N)/c, mida saab leida perpendikulaaride joonestamisel OLEN. Ja M.D. Leitud väärtus (see punkt on joonisel tähistatud tähega D) kaks punkti vastavad R Ja G kõveral (3.3.11). Kasutades neid punkte, joonestades risti, leiame kaks punkti korraga E" Ja E", mis asuvad kõveral (3.3.9). Nende koordinaadid:

Perpendikulaari joonistamine OLEN., ületasime kõvera (3.3.10) veel ühes punktis IN. See punkt vastab samale R Ja K kõveral (3.3.11) ja sama N Ja SCH. Koordineerida N selle punkti saab leida, kukutades risti alla IN telje kohta PEAL. Nii et saame punktid F" ja F", mis asub samuti kõveral (3.3.9).
Tulevad teisest punktist N, samamoodi saame kõveral (3.3.9) asuvad uued neli punkti. Erandiks jääb asja mõte Ni= ?2/72- Selle põhjal saame ainult kaks punkti: TO Ja L. Need on kõvera alumine ja ülemine punkt (3.3.9).
Ei saa lähtuda väärtustest N, ja väärtustest N2 . Suund alates N2 kõverale (3.3.11), siis tõustes sirgele Y = cX ja sealt üle kõvera (3.3.10) leiame ka neli kõvera punkti (3.3.9). Erandiks jääb asja mõte Ei=?1/71- Selle põhjal saame ainult kaks punkti: G Ja TO. Need on kõvera (3.3.9) kõige vasakpoolsemad ja parempoolsemad punktid. Erinevalt küsides N Ja N2 ja olles saanud päris palju punkte, neid ühendades konstrueerime ligikaudselt kõvera (3.3.9).
Konstruktsioonist on selge, et tegemist on suletud kõveraga, mis sisaldab endas punkti 12 = (?2/721?1/71)”, alustades teatud lähteandmetest N Yu ja N20. Võttes teise väärtuse C, st. muud lähteandmed, saame teise suletud kõvera, mis ei ristu esimest ja sisaldab endas ka punkti (?2/721 ?1/71)1. Seega on trajektooride perekond (3.3.9) punkti 12 ümbritsevate suletud joonte perekond (vt joonis 3.3). Uurime selle ainsuse punkti stabiilsuse tüüpi Ljapunovi meetodi abil.
Kuna kõik parameetrid e 1, ?2, 71,72 on positiivsed, punkt (N[ asub faasitasandi positiivses kvadrandis. Süsteemi lineariseerimine selle punkti lähedal annab:

Siin n(t) ja 7i2(N1, N2 :
Süsteemi (3.3.13) iseloomulik võrrand:

Selle võrrandi juured on puhtalt kujuteldavad:

Seega näitab süsteemi uurimine, et ainsuse punkti lähedal olevad trajektoorid on kujutatud kontsentriliste ellipsidega ja ainsuse punkt ise on keskpunkt (joonis 3.4). Vaadeldaval Volterra mudelil on ka suletud trajektoorid ainsuse punktist kaugel, kuigi nende trajektooride kuju on juba erinev ellipsoidsest. Muutujate käitumine Ni, N2 aja jooksul on näidatud joonisel fig. 3.5.

Riis. 3.4.

Riis. 3.5. Saagiarvude sõltuvus N mina ja kiskja N2 ajast
Kesk-tüüpi ainsuse punkt on stabiilne, kuid mitte asümptootiliselt. Näitame selle näitega, mis see on. Las kõhklused Ni(t) ja LGgM esinevad nii, et esituspunkt liigub piki faasitasandit mööda trajektoori 1 (vt joonis 3.4). Hetkel, kui punkt on asendis M, lisatakse süsteemi väljastpoolt teatud arv isendeid N 2, nii et esindav punkt hüppab punktist M punktini A/". Pärast seda, kui süsteem jälle endale jätta, siis võnkumised Ni Ja N2 toimub juba varasemast suuremate amplituudidega ja esituspunkt liigub mööda trajektoori 2. See tähendab, et süsteemi võnkumised on ebastabiilsed: välismõjul muudavad nad igaveseks oma omadusi. Tulevikus käsitleme mudeleid, mis kirjeldavad stabiilseid võnkerežiime ja näitavad, et faasitasandil kujutatakse selliseid asümptootilisi stabiilseid perioodilisi liikumisi piirtsüklite abil.
Joonisel fig. Joonisel 3.6 on näidatud katsekõverad – karusloomade arvu kõikumised Kanadas (Hudson's Bay Company andmetel). Need kõverad on koostatud koristatud nahkade arvu andmete põhjal. Jäneste (saakloomade) ja ilveste (kiskjate) arvukuse kõikumise perioodid on ligikaudu ühesugused ja jäävad suurusjärku 9–10 aastat. Sel juhul on maksimaalne jäneste arv reeglina aasta võrra ees maksimaalsest ilveste arvust.
Nende eksperimentaalsete kõverate kuju on palju vähem korrapärane kui teoreetilised. Kuid sel juhul piisab, kui mudel tagab teoreetilise ja eksperimentaalse kõvera kõige olulisemate karakteristikute kokkulangemise, s.o. amplituudiväärtused ja faasinihked kiskjate ja saaklooma arvukuse kõikumiste vahel. Volterra mudeli märksa tõsisem puudus on võrrandisüsteemi lahenduste ebastabiilsus. Tõepoolest, nagu eespool mainitud, peaks iga juhuslik muutus ühe või teise liigi arvukuses mudelit järgides kaasa tooma mõlema liigi võnkeamplituudi muutumise. Loomulikult puutuvad loomad looduslikes tingimustes kokku lugematute selliste juhuslike mõjudega. Nagu katsekõveratelt näha, on liikide arvukuse kõikumiste amplituud aastate lõikes vähe erinev.
Volterra mudel on matemaatilise ökoloogia võrdlusalus (baas) samal määral, kui harmoonilise ostsillaatori mudel on klassikalise ja kvantmehaanika jaoks põhiline. Selle mudeli kasutamine, mis põhineb väga lihtsustatud ideedel süsteemi käitumist kirjeldavate mustrite olemuse kohta, on puhtalt matemaatiline
3. peatükk

Riis. 3.6. Karusloomade arvukuse kineetilised kõverad Hudson Bay Fur Company andmetel (Seton-Thomson, 1987) Tehti järeldus sellise süsteemi käitumise kvalitatiivse olemuse kohta - karusloomade olemasolu kohta. rahvastiku kõikumised sellises süsteemis. Ilma matemaatilise mudeli konstrueerimata ja seda kasutamata oleks selline järeldus võimatu.
Kõige lihtsamal kujul, mida me eespool käsitlesime, on Volterra süsteemil kaks põhilist ja omavahel seotud puudust. Nende "kõrvaldamisele" on pühendatud ulatuslik ökoloogiline ja matemaatiline kirjandus. Esiteks muudab mis tahes lisategurite kaasamine mudelisse kvalitatiivselt süsteemi käitumist. Mudeli teine "bioloogiline" puudus seisneb selles, et see ei sisalda põhiomadusi, mis on omased ühelegi populatsioonipaarile, kes interakteeruvad röövloom-saagi põhimõttel: kiskja küllastumise mõju, röövlooma ja saagi piiratud ressursid. saagi ülejäägiga, röövloomale saadaoleva minimaalse saagiarvu võimalusega jne.
Nende puuduste kõrvaldamiseks on erinevate autorite poolt välja pakutud erinevaid Volterra süsteemi modifikatsioone. Huvitavamaid neist käsitletakse punktis 3.5. Siin keskendume ainult mudelile, mis võtab arvesse mõlema populatsiooni kasvu enesepiiranguid. Selle mudeli näide näitab selgelt, kuidas otsuste olemus võib süsteemi parameetrite muutumisel muutuda.
Niisiis, me kaalume süsteemi

Süsteem (3.3.15) erineb varem vaadeldud süsteemist (3.3.6) selle poolest, et võrrandite paremal küljel on terminid kujul -7 uNf,
Need terminid peegeldavad tõsiasja, et röövloomade populatsioon ei saa piiramatult kasvada ka röövloomade puudumisel piiratud toiduvarude ja piiratud elupaiga tõttu. Samad "enesepiirangud" on kehtestatud röövloomade populatsioonile.
Liikide statsionaarsete arvude leidmiseks iVi ja N2 Võrdlustame süsteemi (3.3.15) võrrandite parempoolsed küljed nulliga. Lahendused, kus röövloomade või saakloomade arv on null, ei paku meile praegu huvi. Seetõttu kaaluge algebralist süsteemi
võrrandid  Tema otsus
Tema otsus
annab meile ainsuse punkti koordinaadid. Siin tuleks süsteemi parameetritele kehtestada statsionaarsete numbrite positiivsuse tingimus: N> 0 ja N2 > 0. Ainsuse punkti (3.3.16) naabruses lineariseeritud süsteemi tunnusvõrrandi juured:

Iseloomulike arvude avaldisest on selge, et kui tingimus on täidetud
siis kiskjate ja saaklooma arvukus läbib aja jooksul summutatud võnkumisi, süsteemil on nullist erinev ainsuspunkt ja stabiilne fookus. Sellise süsteemi faasiportree on näidatud joonisel fig. 3,7 a.
Oletame, et ebavõrdsuse (3.3.17) parameetrid muudavad oma väärtusi nii, et tingimus (3.3.17) muutub võrdsuseks. Siis on süsteemi (3.3.15) iseloomulikud arvud võrdsed ja selle ainsuse punkt asub stabiilsete fookuste ja sõlmede piirkondade piiril. Kui ebavõrdsuse märk (3.3.17) pöörata ümber, muutub ainsuse punkt stabiilseks sõlmeks. Süsteemi faasiportree selle juhtumi jaoks on näidatud joonisel fig. 3.76.
Nagu ühe üldkogumi puhul, saab mudeli (3.3.6) jaoks välja töötada stohhastilise mudeli, kuid eksplitsiitset lahendust sellele ei saa. Seetõttu piirdume üldiste kaalutlustega. Oletame näiteks, et tasakaalupunkt asub igast teljest teatud kaugusel. Seejärel faasitrajektooride puhul, millel JVj väärtused, N2 piisavalt suureks, on deterministlik mudel üsna rahuldav. Aga kui ühel hetkel


Riis. 3.7. Süsteemi faasiportree (3.3.15): A - kui parameetrite vaheline seos (3.3.17) on täidetud; b- parameetrite vahelise pöördsuhte teostamisel
faasitrajektoori korral ei ole ükski muutuja väga suur, siis võivad juhuslikud kõikumised muutuda oluliseks. Need viivad selleni, et esinduspunkt liigub ühele teljele, mis tähendab vastava liigi väljasuremist. Seega osutub stohhastiline mudel ebastabiilseks, kuna stohhastiline “triiv” viib varem või hiljem ühe liigi väljasuremiseni. Seda tüüpi mudeli puhul sureb kiskja lõpuks välja kas juhuslikult või seetõttu, et tema saagipopulatsioon likvideeritakse kõigepealt. Kiskja-saakloomade süsteemi stohhastiline mudel seletab hästi Gause'i katseid (Gause, 1934; 2000), kus ripsloomad Paramettum candatum oli ühe teise ripslase ohvriks Didinium nasatum- kiskja. Deterministlike võrrandite (3.3.6) järgi eeldati nendes katsetes ligikaudu viis isendit igast liigist, mistõttu pole üllatav, et iga korduva katse puhul surid kas kiskjad või saakloomad üsna kiiresti välja (ja seejärel kiskjad ). .
Seega näitab Volterra liikide interaktsiooni mudelite analüüs, et vaatamata selliste süsteemide käitumistüüpide mitmekesisusele, ei saa konkureerivate liikide mudelis üldse esineda arvukuses sumbutamata kõikumisi. Kiskja-saagi mudelis ilmnevad summutamata võnkumised tänu mudeli võrrandite (3.3.6) erikuju valikule. Sel juhul muutub mudel ebakareseks, mis näitab, et sellises süsteemis puuduvad mehhanismid, mis püüavad oma olekut säilitada. Selliseid võnkumisi täheldatakse aga looduses ja katsetes. Vajadus nende teoreetilise seletuse järele oli üheks põhjuseks mudelikirjelduste üldisemal kujul formuleerimiseks. Jaotis 3.5 on pühendatud selliste üldiste mudelite käsitlemisele.