Matematisk modell konservativt rovdyrbytte. Kurs: Kvalitativ studie av rovdyr-byttedyr-modellen. Simuleringsmodellering av "Predator-Prey"-systemet
Rovdyr kan spise planteetere, og også svake rovdyr. Rovdyr har et bredt utvalg av mat, bytter enkelt fra ett bytte til et annet, mer tilgjengelig. Rovdyr angriper ofte svake byttedyr. En økologisk balanse opprettholdes mellom byttedyr-rovdyrpopulasjoner.[ ...]
Hvis likevekten er ustabil (det er ingen grensesykluser) eller den ytre syklusen er ustabil, vil tallene for begge artene, som opplever sterke svingninger, forlate likevektens nærhet. Dessuten oppstår rask degenerasjon (i den første situasjonen) med lav tilpasning av rovdyret, dvs. med sin høye dødelighet (sammenlignet med reproduksjonshastigheten til offeret). Dette betyr at et rovdyr som er svakt på alle måter ikke bidrar til stabilisering av systemet og dør ut av seg selv.[ ...]
Predatorpresset er spesielt sterkt når, i samevolusjon mellom rovdyr og byttedyr, balansen skifter mot rovdyret og byttets rekkevidde blir smalere. Konkurransekamp er nært knyttet til mangel på matressurser, det kan også være en direkte kamp for eksempel av rovdyr om plass som ressurs, men som oftest er det rett og slett fortrengning av en art som ikke har nok mat i en gitt territorium av en art som har nok av samme mengde mat. Dette er konkurranse mellom arter.[ ...]
 |
Til slutt, i "rovdyr-byttedyr"-systemet beskrevet av modell (2.7), er forekomsten av diffusjonsustabilitet (med lokal likevektsstabilitet) bare mulig hvis den naturlige dødeligheten til rovdyret øker med populasjonen raskere enn den lineære funksjonen, og trofisk funksjon skiller seg fra Volterra eller når byttedyrpopulasjonen er en populasjon av Ollie-typen.[ ...]
Teoretisk sett, i "ett rovdyr - to byttedyr"-modeller, kan ekvivalent predasjon (manglende preferanse for en eller annen type byttedyr) påvirke den konkurransedyktige sameksistensen av byttedyrarter bare på de stedene hvor en potensielt stabil likevekt allerede eksisterer. Mangfoldet kan bare øke under forhold der arter med mindre konkurranseevne har høyere bestandsvekst enn dominerende arter. Dette gjør det mulig å forstå situasjonen når selv beite fører til økt planteartsmangfold der et større antall arter som er valgt ut for rask reproduksjon sameksisterer med arter hvis utvikling er rettet mot å øke konkurranseevnen.[ ...]
På samme måte kan valget av bytte, avhengig av dens tetthet, føre til en stabil likevekt i teoretiske modeller av to konkurrerende byttetyper, hvor det ikke eksisterte likevekt før. For å gjøre dette, må rovdyret være i stand til funksjonelle og numeriske reaksjoner på endringer i byttedyrtetthet; det er imidlertid mulig at bytte (uforholdsmessig hyppige angrep på det mest utbredte offeret) vil være viktigere i dette tilfellet. Det er faktisk funnet at bytte har en stabiliserende effekt i "one predator - n prey"-systemer og er den eneste mekanismen som er i stand til å stabilisere interaksjoner i tilfeller der byttedyrnisjene overlapper fullstendig. Denne rollen kan spilles av uspesialiserte rovdyr. Preferansen til mer spesialiserte rovdyr for en dominerende konkurrent virker på samme måte som bytte av rovdyr, og kan stabilisere teoretiske interaksjoner i modeller der det ikke var likevekt mellom byttedyr før, forutsatt at deres nisjer til en viss grad er atskilt.[ .. .]
Fellesskapet er heller ikke stabilisert og rovdyret er 'sterkt på alle måter', dvs. godt tilpasset et gitt byttedyr og med lav relativ dødelighet. I dette tilfellet har systemet en ustabil grensesyklus og, til tross for stabiliteten til likevektsposisjonen, degenererer det i et tilfeldig miljø (rovdyret spiser bort byttet og dør som et resultat). Denne situasjonen tilsvarer langsom degenerasjon.[ ...]
Dermed kan det med en god tilpasning av et rovdyr i nærheten av en stabil likevekt oppstå ustabile og stabile sykluser, d.v.s. avhengig av startforholdene, har "rovdyr-byttedyr"-systemet enten en tendens til likevekt, eller svingende, forlater det, eller stabile svingninger i antallet av begge arter etableres i nærheten av likevekten.[ ...]
Organismer som er klassifisert som rovdyr lever av andre organismer og ødelegger byttet deres. Blant levende organismer bør det derfor skilles ut ett mer klassifiseringssystem, nemlig "rovdyr" og "ofre". Forholdet mellom slike organismer har utviklet seg gjennom hele utviklingen av livet på planeten vår. Rovorganismer fungerer som naturlige regulatorer av antall byttedyr. En økning i antall "rovdyr" fører til en nedgang i antall "byttedyr", som igjen reduserer tilgangen på mat ("byttedyr") for "rovdyrene", som generelt tilsier en nedgang i antallet av "byttedyr" osv. I In the biocenosis er det altså konstante svingninger i antall rovdyr og byttedyr, generelt etableres en viss balanse i en viss tidsperiode innenfor ganske stabile miljøforhold.[ ... ]
Dette kommer til slutt til en økologisk balanse mellom rovdyr- og byttedyrpopulasjoner.[ ...]
For en trofisk funksjon av den tredje typen vil likevektstilstanden være stabil hvis hvor N er funksjonens bøyningspunkt (se fig. 2, c). Dette følger av det faktum at den trofiske funksjonen er konkav i intervallet, og følgelig øker den relative andelen av byttedyrforbruket til rovdyret.[ ...]
La Гг = -Г, dvs. det er et fellesskap av typen "rovdyr-byttedyr". I dette tilfellet er første ledd i uttrykk (7.4) lik null, og for å oppfylle vilkåret om stabilitet med hensyn til sannsynligheten for likevektstilstanden N kreves det at det andre leddet heller ikke er positivt.[ ...]
For det betraktede fellesskapet av rovdyr-byttetypen kan vi derfor konkludere med at den generelt positive likevekten er asymptotisk stabil, dvs. for alle innledende data forutsatt at N >0.[ ...]
Så, i et homogent miljø som ikke har tilfluktsrom for reproduksjon, ødelegger et rovdyr før eller siden byttedyrbestanden og dør deretter ut av seg selv. Waves of life» (endringer i antall rovdyr og byttedyr) følger hverandre med et konstant faseskifte, og i gjennomsnitt forblir antallet både rovdyr og byttedyr omtrent på samme nivå. Varigheten av perioden avhenger av vekstratene til begge artene og av de innledende parameterne. For byttedyrpopulasjonen er påvirkningen av rovdyret positiv, siden dens overdrevne reproduksjon ville føre til kollaps av antallet. På sin side bidrar alle mekanismene som forhindrer fullstendig utryddelse av byttet til bevaring av rovdyrets næringsgrunnlag.[ ...]
Andre modifikasjoner kan skyldes oppførselen til rovdyret. Antall byttedyr som et rovdyr er i stand til å konsumere på et gitt tidspunkt har sin grense. Effekten av metning av rovdyret når man nærmer seg denne grensen er vist i tabell. 2-4, B. Interaksjonene beskrevet av ligning 5 og 6 kan ha stabile likevektspunkter eller utvise sykliske svingninger. Imidlertid skiller slike sykluser seg fra de som reflekteres i Lotka-Volterra-ligningene 1 og 2. Syklusene som formidles av ligningene 5 og 6 kan ha konstant amplitude og gjennomsnittlig tetthet så lenge mediet er konstant; etter at et brudd har skjedd, kan de gå tilbake til sine tidligere amplituder og gjennomsnittlige tettheter. Slike sykluser, som gjenopprettes etter brudd, kalles stabile grensesykluser. Samspillet mellom en hare og en gaupe kan betraktes som en stabil grensesyklus, men dette er ikke en Lotka-Volterra-syklus.[ ...]
La oss vurdere forekomsten av diffusjonsustabilitet i "rovdyr-byttedyr"-systemet, men først skriver vi ut forholdene som sikrer forekomsten av diffusjonsustabilitet i systemet (1.1) ved n = 2. Det er klart at likevekten (N , W) er lokal (dvs. [ .. .]
La oss vende oss til tolkningen av saker knyttet til den langsiktige sameksistensen av rovdyr og byttedyr. Det er klart at i fravær av grensesykluser vil en stabil likevekt tilsvare populasjonssvingninger i et tilfeldig miljø, og deres amplitude vil være proporsjonal med spredningen av forstyrrelser. Et slikt fenomen vil oppstå dersom rovdyret har høy relativ dødelighet og samtidig høy grad av tilpasning til et gitt bytte.[ ...]
La oss nå vurdere hvordan dynamikken til systemet endres med en økning i rovdyrets kondisjon, dvs. med synkende b fra 1 til 0. Hvis kondisjonen er lav nok, er det ingen grensesykluser, og likevekten er ustabil. Med veksten av kondisjon i nærheten av denne likevekten, er fremveksten av en stabil syklus og deretter en ekstern ustabil en mulig. Avhengig av startforholdene (forholdet mellom rovdyr- og byttedyrbiomasse) kan systemet enten miste stabilitet, dvs. forlate likevektsområdet, eller stabile oscillasjoner vil bli etablert i det over tid. Ytterligere vekst av kondisjon gjør den oscillerende karakteren til systemets oppførsel umulig. Men når b [...]
Et eksempel på negativ (stabiliserende) tilbakemelding er forholdet mellom rovdyr og byttedyr eller funksjonen til havkarbonatsystemet (løsning av CO2 i vann: CO2 + H2O -> H2CO3). Normalt er mengden karbondioksid oppløst i havvann i delvis likevekt med konsentrasjonen av karbondioksid i atmosfæren. Lokale økninger i karbondioksid i atmosfæren etter vulkanutbrudd fører til intensivering av fotosyntesen og dens absorpsjon av karbonatsystemet i havet. Når nivået av karbondioksid i atmosfæren synker, frigjør karbonatsystemet i havet CO2 til atmosfæren. Derfor er konsentrasjonen av karbondioksid i atmosfæren ganske stabil.[ ...]
[ ...]
Som R. Ricklefs (1979) bemerker, er det faktorer som bidrar til stabilisering av relasjoner i "rovdyr-byttedyr"-systemet: rovdyrets ineffektivitet, tilstedeværelsen av alternative matressurser i rovdyret, en reduksjon i forsinkelsen i rovdyrets reaksjon, samt miljørestriksjoner som det ytre miljø pålegger en eller flere forskjellige populasjoner. Interaksjoner mellom rovdyr- og byttedyrpopulasjoner er svært mangfoldige og komplekse. Således, hvis rovdyr er effektive nok, kan de regulere tettheten til byttedyrpopulasjonen, og holde den på et nivå under miljøets kapasitet. Gjennom påvirkningen de har på byttedyrpopulasjoner, påvirker rovdyr utviklingen av ulike byttedyregenskaper, noe som til slutt fører til en økologisk balanse mellom rovdyr- og byttedyrpopulasjoner.[ ...]
Dersom ett av vilkårene er oppfylt: 0 1/2. Hvis 6 > 1 (kA [ ...]
Stabiliteten til biotaen og miljøet avhenger bare av samspillet mellom planter - autotrofer og planteetende heterotrofe organismer. Rovdyr av enhver størrelse er ikke i stand til å forstyrre den økologiske balansen i samfunnet, siden de under naturlige forhold ikke kan øke antallet med et konstant antall byttedyr. Rovdyr må ikke bare være i bevegelse selv, men kan bare spise av dyr i bevegelse.[ ...]
Ingen annen fisk er så utbredt som gjedder. I de få fiskeområdene i stillestående eller rennende vann er det ikke noe press fra gjedde for å opprettholde balansen mellom byttedyr og rovdyr. Gjedde er usedvanlig godt representert i verden. De fanges over hele den nordlige halvkule fra USA og Canada i Nord-Amerika, gjennom Europa til Nord-Asia.[ ...]
En annen mulighet for stabil sameksistens oppstår her, i et snevert område med relativt høy tilpasning. Ved overgang til et ustabilt regime med et veldig "godt" rovdyr, kan det oppstå en stabil ytre begrensende syklus, der spredningen av biomasse balanseres av dens tilstrømning til systemet (byttets høye produktivitet). Så oppstår en merkelig situasjon når det mest sannsynlige er to karakteristiske verdier av amplituden til tilfeldige svingninger. Noen forekommer nær likevekt, andre nær grensesyklusen, og mer eller mindre hyppige overganger mellom disse modusene er mulig.[ ...]
Hypotetiske populasjoner som oppfører seg i henhold til vektorene i fig. 10.11 A, vist i fig. 10.11,-B ved hjelp av en graf som viser dynamikken i forholdet mellom antall rovdyr og byttedyr og i fig. 10.11.5 i form av en graf over dynamikken til antall rovdyr og byttedyr over tid. I byttedyrpopulasjonen, når den beveger seg fra en likevekt med lav tetthet til en likevekt med høy tetthet og vender tilbake, oppstår det et "flash" av tall. Og dette utbruddet er ikke et resultat av en like uttalt endring i miljøet. Tvert imot, denne endringen i antall genereres av selve påvirkningen (med et lavt nivå av "støy" i miljøet), og spesielt gjenspeiler den eksistensen av flere likevektstilstander. Lignende resonnement kan brukes for å forklare mer komplekse tilfeller av populasjonsdynamikk i naturlige populasjoner.[ ...]
Den viktigste egenskapen til et økosystem er dets stabilitet, utvekslingsbalansen og prosessene som skjer i det. Befolkningers eller økosystemers evne til å opprettholde en stabil dynamisk balanse i skiftende miljøforhold kalles homeostase (homoios - det samme, lignende; stasis - tilstand). Homeostase er basert på prinsippet om tilbakemelding. For å opprettholde balansen i naturen er det ikke nødvendig med ekstern kontroll. Et eksempel på homeostase er "rovdyr-byttedyr"-subsystemet, der tettheten av rovdyr- og byttedyrpopulasjoner er regulert.[ ...]
Et naturlig økosystem (biogeocenose) fungerer stabilt med den konstante interaksjonen mellom elementene, sirkulasjonen av stoffer, overføring av kjemisk, energi, genetisk og annen energi og informasjon gjennom kjedekanaler. I henhold til prinsippet om likevekt har ethvert naturlig system med en strøm av energi og informasjon som passerer gjennom det en tendens til å utvikle en stabil tilstand. Samtidig tilveiebringes stabiliteten til økosystemene automatisk på grunn av tilbakemeldingsmekanismen. Tilbakemelding består i å bruke dataene mottatt fra de administrerte komponentene i økosystemet for å gjøre justeringer av styringskomponentene i prosessen. Forholdet "rovdyr" - "bytte" omtalt ovenfor i denne sammenheng kan beskrives noe mer detaljert; så i det akvatiske økosystemet spiser rovfisk (gjedde i dammen) andre typer byttefisk (krysskarpe); hvis antall karpe vil øke, er dette et eksempel på positive tilbakemeldinger; gjedde, som fôrer på crucian karpe, reduserer antallet - dette er et eksempel på negativ tilbakemelding; med en økning i antall rovdyr, reduseres antallet ofre, og rovdyret, som mangler mat, reduserer også veksten av befolkningen; til slutt, i dammen som vurderes, etableres en dynamisk balanse i mengden av både gjedde og karpe. En balanse opprettholdes hele tiden som vil utelukke forsvinningen av ethvert ledd i trofiskkjeden (fig. 64).[ ...]
La oss gå videre til den viktigste generaliseringen, nemlig at negative interaksjoner blir mindre merkbare over tid hvis økosystemet er tilstrekkelig stabilt og dets romlige struktur tillater gjensidig tilpasning av bestander. I modellsystemer av rovdyr-byttedyr-typen, beskrevet av Lotka-Volterra-ligningen, hvis ytterligere termer ikke er introdusert i ligningen som karakteriserer effekten av faktorer for selvbegrensning av populasjonen, oppstår fluktuasjoner kontinuerlig og dør ikke ut (se Levontin, 1969). Pimentel (1968; se også Pimentel og Stone, 1968) viste eksperimentelt at slike tilleggsbegreper kan reflektere gjensidige tilpasninger eller genetisk tilbakemelding. Da nye kulturer ble skapt fra individer som tidligere hadde eksistert i en kultur i to år, hvor antallet var utsatt for betydelige svingninger, viste det seg at de utviklet en økologisk homeostase, der hver av populasjonene ble "undertrykt" av den andre i en slik grad at det viste seg deres sameksistens i en mer stabil likevekt.
Kolmogorovs modell gjør en betydelig antagelse: siden det antas at dette betyr at det er mekanismer i byttedyrpopulasjonen som regulerer deres overflod selv i fravær av rovdyr.
Dessverre tillater ikke en slik formulering av modellen oss å svare på spørsmålet som det har vært mye kontrovers rundt i det siste og som vi allerede nevnte i begynnelsen av kapittelet: hvordan kan en rovdyrbestand utøve en regulerende innflytelse på et byttedyr. befolkning slik at hele systemet er stabilt? Derfor vil vi gå tilbake til modell (2.1), der det ikke finnes selvreguleringsmekanismer (for eksempel regulering gjennom intraspesifikk konkurranse) i byttedyrbestanden (så vel som i rovdyrbestanden); derfor er den eneste mekanismen for å regulere overflod av arter i et samfunn trofiske forhold mellom rovdyr og byttedyr.
Her (så, i motsetning til den forrige modellen, er det naturlig at løsninger (2.1) avhenger av den spesifikke typen trofisk funksjon, som igjen bestemmes av predasjonens natur, dvs. rovdyrets trofiske strategi og byttets forsvarsstrategi Felles for alle disse funksjonene (se fig. I) er følgende egenskaper:
System (2.1) har ett ikke-trivielt stasjonært punkt hvis koordinater bestemmes fra ligningene
![]()
med naturlig begrensning.
Det er ett stasjonært punkt til (0, 0) som tilsvarer trivielle likevekten. Det er lett å vise at dette punktet er en sal, og koordinataksene er separatriser.
Den karakteristiske ligningen for et punkt har formen
![]()
Tydeligvis for den klassiske Volterra-modellen.
Derfor kan verdien av f betraktes som et mål på avviket til den betraktede modellen fra Volterra-modellen.
![]()
det stasjonære punktet er fokus, og oscillasjoner vises i systemet; når den motsatte ulikheten er oppfylt, er det en node, og det er ingen svingninger i systemet. Stabiliteten til denne likevektstilstanden bestemmes av tilstanden
det vil si at det i hovedsak avhenger av typen trofisk funksjon til rovdyret.
Tilstand (5.5) kan tolkes som følger: for stabiliteten av den ikke-trivielle likevekten til rovdyr-bytte-systemet (og dermed for eksistensen av dette systemet), er det tilstrekkelig at i nærheten av denne tilstanden er den relative andelen byttedyr som konsumeres av rovdyret øker med økningen i antall byttedyr. Faktisk er andelen byttedyr (av deres totale antall) konsumert av et rovdyr beskrevet av en differensierbar funksjon hvis veksttilstand (positiviteten til derivatet) ser ut som
![]()
Den siste betingelsen, tatt på punktet, er ikke annet enn betingelsen (5.5) for likevektsstabilitet. Med kontinuitet må det også holde i et eller annet nabolag av punktet. Derfor, hvis antallet ofre i dette nabolaget, så
La nå den trofiske funksjonen V ha formen vist i fig. 11a (karakteristisk for virvelløse dyr). Det kan vises at for alle endelige verdier (siden den er konveks oppover)
dvs. ulikhet (5,5) er ikke tilfredsstilt for noen verdier av det stasjonære antallet ofre.
Dette betyr at i et system med denne typen trofisk funksjon er det ingen stabil ikke-triviell likevekt. Flere utfall er mulige: enten øker antallet av både byttet og rovdyret i det uendelige, eller (når banen passerer nær en av koordinataksene), på grunn av tilfeldige årsaker, vil antallet på byttedyret eller rovdyrets nummer bli. lik null. Hvis byttet dør, vil rovdyret dø etter en tid, men hvis rovdyret dør først, vil antallet byttedyr begynne å øke eksponentielt. Det tredje alternativet - fremveksten av en stabil grensesyklus - er umulig, noe som lett kan bevises.
Faktisk uttrykket

i den positive kvadranten er alltid positiv, med mindre den har formen vist i fig. 11, a. Da er det ifølge Dulacs kriterium ingen lukkede baner i dette området og en stabil grensesyklus kan ikke eksistere.
Så vi kan konkludere: hvis den trofiske funksjonen har formen vist i fig. 11a, så kan ikke rovdyret være en regulator som sikrer stabiliteten til byttedyrbestanden og dermed stabiliteten til hele systemet som helhet. Systemet kan bare være stabilt dersom byttedyrbestanden har sine egne interne reguleringsmekanismer, som intraspesifikk konkurranse eller epizootier. Denne reguleringsmulighet er allerede vurdert i §§ 3, 4.
Det ble tidligere bemerket at denne typen trofisk funksjon er karakteristisk for insektrovdyr, hvis "ofre" også vanligvis er insekter. På den annen side viser observasjoner av dynamikken til mange naturlige samfunn av typen «rovdyr-byttedyr», inkludert insektarter, at de er preget av svingninger med veldig stor amplitude og en veldig spesifikk type.
Vanligvis, etter en mer eller mindre gradvis økning i antallet (som kan forekomme enten monotont eller i form av fluktuasjoner med økende amplitude), oppstår dets skarpe fall (fig. 14), og deretter gjentar mønsteret seg selv. Tilsynelatende kan denne arten av dynamikken til overflod av insektarter forklares av ustabiliteten til dette systemet ved lave og middels verdier av overflod og virkningen av kraftige intrapopulasjonsregulatorer av overflod ved store verdier.

Ris. Fig. 14. Populasjonsdynamikk til den australske psylliden Cardiaspina albitextura som lever av eukalypter. (Fra artikkelen: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, nr. 3, s. 362-380.)
Hvis "rovdyr-byttedyr"-systemet inkluderer arter som er i stand til ganske kompleks oppførsel (for eksempel er rovdyr i stand til å lære eller byttedyr er i stand til å finne ly), så kan en stabil ikke-triviell likevekt eksistere i et slikt system. Beviset for denne påstanden er ganske enkelt.
Den trofiske funksjonen bør da ha formen vist i fig. 11, c. Punktet på denne grafen er kontaktpunktet for den rette linjen trukket fra origo for koordinater med grafen til den trofiske funksjonen. Det er åpenbart at funksjonen på dette punktet har et maksimum. Det er også lett å vise at betingelsen (5.5) er oppfylt for alle. Derfor vil en ikke-triviell likevekt der antall ofre er mindre være asymptotisk stabil
Vi kan imidlertid ikke si noe om hvor stor stabilitetsregionen til denne likevekten er. For eksempel, hvis det er en ustabil grensesyklus, må denne regionen ligge inne i syklusen. Eller en annen variant: den ikke-trivielle likevekten (5.2) er ustabil, men det er en stabil grensesyklus; i dette tilfellet kan man også snakke om stabiliteten til rovdyr-bytte-systemet. Siden uttrykk (5.7) når du velger en trofisk funksjon som fig. 11, i kan endre fortegn ved endring ved , så fungerer ikke Dulac-kriteriet her og spørsmålet om eksistensen av grensesykluser forblir åpent.

Interaksjon mellom individer i "rovdyr-byttedyr"-systemet
5. års elev 51 A gruppe
Institutt for bioøkologi
Nazarova A.A.
Vitenskapelig rådgiver:
Podshivalov A.A.
Orenburg 2011
INTRODUKSJON
INTRODUKSJON
I våre daglige resonnementer og observasjoner blir vi, uten å vite det selv, og ofte uten å være klar over det, styrt av lover og ideer som ble oppdaget for mange tiår siden. Med tanke på rovdyr-bytte-problemet, tipper vi at byttet også indirekte påvirker rovdyret. Hva ville en løve spist hvis det ikke fantes antiloper; hva ville ledere gjort hvis det ikke fantes arbeidere; hvordan utvikle en virksomhet hvis kundene ikke har midler ...
"Rovdyr-byttedyr"-systemet er et komplekst økosystem der langsiktige forhold mellom rovdyr og byttedyr er realisert, et typisk eksempel på samevolusjon. Forholdet mellom rovdyr og byttet deres utvikler seg syklisk, og er en illustrasjon på en nøytral likevekt.
Studiet av denne formen for relasjoner mellom arter, i tillegg til å oppnå interessante vitenskapelige resultater, lar oss løse mange praktiske problemer:
optimalisering av biotekniske tiltak både i forhold til byttearter og i forhold til rovdyr;
forbedre kvaliteten på territoriell beskyttelse;
regulering av jaktpress i fangstbruk mv.
Det foregående bestemmer relevansen til det valgte emnet.
Formålet med kursarbeidet er å studere interaksjonen mellom individer i "rovdyr - byttedyr"-systemet. For å nå målet ble følgende oppgaver satt:
predasjon og dens rolle i dannelsen av trofiske relasjoner;
hovedmodellene for forholdet "rovdyr - byttedyr";
påvirkningen av den sosiale livsstilen i stabiliteten til "rovdyr-byttedyr"-systemet;
laboratoriemodellering av "rovdyr - byttedyr"-systemet.
Påvirkningen av rovdyr på antall byttedyr og omvendt er ganske åpenbar, men det er ganske vanskelig å bestemme mekanismen og essensen av denne interaksjonen. Disse spørsmålene har jeg tenkt å ta opp i kursarbeidet.
#�������######################################## ######"#5#@#?#8#;#0###��####################+##### ######��\############## ###############��#���##### ######## Kapittel 4KAPITTEL 4. LABORATORIEMODELLER AV ROVDYRET - BYTTESYSTEMET
Forskere fra Duke University, i samarbeid med kolleger fra Stanford University, Howard Hughes Medical Institute og California Institute of Technology, som arbeider under ledelse av Dr. Lingchong You, har utviklet et levende system av genmodifiserte bakterier som vil tillate mer detaljerte studier av rovdyr-byttedyr-interaksjoner på et populasjonsnivå.
Den nye eksperimentelle modellen er et eksempel på et kunstig økosystem der forskere programmerer bakterier til å utføre nye funksjoner for å skape. Slike omprogrammerte bakterier kan bli mye brukt i medisin, miljøopprydding og utvikling av biodatamaskiner. Som en del av dette arbeidet omskrev forskerne "programvaren" til E. coli (Escherichia coli) på en slik måte at to forskjellige bakteriepopulasjoner i laboratoriet dannet et typisk system av rovdyr-byttedyr-interaksjoner, et trekk ved dette var at bakteriene slukte ikke hverandre, men kontrollerte antallet motstanderbefolkningen ved å endre frekvensen av "selvmord".
Forskningsfeltet kjent som syntetisk biologi dukket opp rundt 2000, og de fleste systemene som er opprettet siden den gang har vært basert på omprogrammering av en enkelt bakterie. Modellen utviklet av forfatterne er unik ved at den består av to bakteriepopulasjoner som lever i samme økosystem, hvis overlevelse avhenger av hverandre.
Nøkkelen til en vellykket funksjon av et slikt system er to populasjoners evne til å samhandle med hverandre. Forfatterne skapte to stammer av bakterier - "rovdyr" og "planteetere", avhengig av situasjonen, og frigjorde giftige eller beskyttende forbindelser til det generelle økosystemet.
Prinsippet for drift av systemet er basert på å opprettholde forholdet mellom antall rovdyr og byttedyr i et regulert miljø. Endringer i antall celler i en av populasjonene aktiverer omprogrammerte gener, som utløser syntesen av visse kjemiske forbindelser.
Dermed forårsaker et lite antall ofre i miljøet aktivering av selvdestruksjonsgenet i rovdyrceller og deres død. Men etter hvert som antallet ofre øker, når forbindelsen som frigjøres av dem til miljøet en kritisk konsentrasjon og aktiverer rovdyrgenet, som sikrer syntesen av en "motgift" mot selvmordsgenet. Dette fører til en økning i bestanden av rovdyr, som igjen fører til akkumulering av en forbindelse syntetisert av rovdyr i miljøet, og presser ofrene til å begå selvmord.
Ved hjelp av fluorescensmikroskopi dokumenterte forskere interaksjoner mellom rovdyr og byttedyr.
Predatorceller, farget grønt, forårsaker selvmord av byttedyrceller, farget rødt. Forlengelse og brudd på offercellen indikerer dens død.
Dette systemet er ikke en nøyaktig representasjon av rovdyr-byttedyr-interaksjoner i naturen, som rovdyrbakterier lever ikke av byttedyrbakterier og begge populasjoner konkurrerer om de samme matressursene. Forfatterne mener imidlertid at systemet de har utviklet er et nyttig verktøy for biologisk forskning.
Det nye systemet demonstrerer en klar sammenheng mellom genetikk og populasjonsdynamikk, som i fremtiden vil hjelpe i studiet av påvirkningen av molekylære interaksjoner på populasjonsendring, som er et sentralt tema innen økologi. Systemet gir praktisk talt ubegrensede muligheter for å modifisere variabler for å studere i detalj interaksjonene mellom miljø, genregulering og populasjonsdynamikk.
Dermed, ved å kontrollere det genetiske apparatet til bakterier, er det mulig å simulere prosessene for utvikling og interaksjon av mer komplekse organismer.
KAPITTEL 3
KAPITTEL 3
Økologer fra USA og Canada har vist at gruppelivsstilen til rovdyr og deres byttedyr radikalt endrer oppførselen til rovdyr-bytte-systemet og gjør det mer motstandsdyktig. Denne effekten, bekreftet av observasjoner av dynamikken til antall løver og gnuer i Serengeti-parken, er basert på det enkle faktum at med en gruppelivsstil reduseres hyppigheten av tilfeldige møter mellom rovdyr og potensielle ofre.
Økologer har utviklet en rekke matematiske modeller som beskriver oppførselen til rovdyr-bytte-systemet. Spesielt disse modellene forklarer godt de observerte til tider konsistente periodiske svingningene i mengden av rovdyr og byttedyr.
Slike modeller er vanligvis preget av et høyt nivå av ustabilitet. Med andre ord, med et bredt spekter av inngangsparametere (som dødelighet av rovdyr, effektivitet av konvertering av byttedyrbiomasse til rovdyrbiomasse, etc.) i disse modellene, dør før eller siden alle rovdyr enten ut eller spiser først alt byttet, og så fortsatt dø av sult.
I naturlige økosystemer er selvfølgelig alt mer komplisert enn i en matematisk modell. Tilsynelatende er det mange faktorer som kan øke stabiliteten til rovdyr-bytte-systemet, og i realiteten kommer det sjelden til så skarpe hopp i antall som hos Canada gaupe og hare.
Økologer fra Canada og USA publiserte i siste utgave av tidsskriftet " natur" en artikkel som trakk oppmerksomheten til én enkel og åpenbar faktor som dramatisk kan endre oppførselen til rovdyr-bytte-systemet. Det handler om gruppeliv.
De fleste tilgjengelige modeller er basert på antakelsen om en jevn fordeling av rovdyr og deres byttedyr innenfor et gitt territorium. Dette er grunnlaget for å beregne hyppigheten av deres møter. Det er klart at jo høyere tetthet av byttedyr, jo oftere snubler rovdyr over dem. Antall angrep, inkludert vellykkede, og til syvende og sist intensiteten av predatorer avhenger av dette. For eksempel, med et overskudd av byttedyr (hvis du ikke trenger å bruke tid på å søke), vil spisehastigheten bare begrenses av tiden det er nødvendig for rovdyret å fange, drepe, spise og fordøye det neste byttet. Hvis byttet sjelden fanges, blir hovedfaktoren som bestemmer beitehastigheten tiden det tar å lete etter byttet.
I de økologiske modellene som brukes for å beskrive "rovdyr-byttedyr"-systemene, spiller arten av avhengigheten til predasjonsintensiteten (antall byttedyr spist av ett rovdyr per tidsenhet) av byttedyrbestandstettheten en nøkkelrolle. Sistnevnte er beregnet som antall dyr per arealenhet.
Det skal bemerkes at med en gruppelivsstil av både byttedyr og rovdyr, er den første antakelsen om en jevn romlig fordeling av dyr ikke tilfredsstilt, og derfor blir alle videre beregninger feil. For eksempel, med en flokklivsstil av byttedyr, vil sannsynligheten for å møte et rovdyr faktisk ikke avhenge av antall individuelle dyr per kvadratkilometer, men av antall flokker per arealenhet. Hvis byttet ble fordelt jevnt, ville rovdyr snuble over dem mye oftere enn i flokkens livsstil, siden det dannes store rom mellom flokkene der det ikke er byttedyr. Et lignende resultat oppnås med gruppens levemåte for rovdyr. En stolthet av løver som vandrer over savannen vil legge merke til få flere potensielle ofre enn en ensom løve som følger samme vei ville gjort.
I tre år (fra 2003 til 2007) utførte forskere nøye observasjoner av løver og deres ofre (først og fremst gnuer) i det enorme territoriet til Serengeti-parken (Tanzania). Befolkningstetthet ble registrert månedlig; intensiteten av å spise av løver av forskjellige arter av hovdyr ble også jevnlig vurdert. Både løvene selv og de syv hovedartene av byttedyrene deres fører en gruppelivsstil. Forfatterne introduserte de nødvendige endringene til standard økologiske formler for å ta hensyn til denne omstendigheten. Parametriseringen av modellene ble utført på grunnlag av reelle kvantitative data innhentet i løpet av observasjoner. Fire versjoner av modellen ble vurdert: i den første ble gruppens levemåte for rovdyr og byttedyr ignorert; i den andre ble den tatt i betraktning bare for rovdyr; i den tredje bare for byttedyr; og i den fjerde, for begge.
|
|
Som man kunne forvente, samsvarte det fjerde alternativet best med virkeligheten. Han viste seg også å være den mest robuste. Dette betyr at med et bredt spekter av inngangsparametere i denne modellen, er langsiktig stabil sameksistens av rovdyr og byttedyr mulig. Dataene fra langtidsobservasjoner viser at modellen også i denne forbindelse reflekterer virkeligheten tilstrekkelig. Antallet løver og deres byttedyr i Serengeti er ganske stabilt, ingenting som ligner periodiske koordinerte svingninger (som tilfellet er med gauper og harer) er observert.
Resultatene som ble oppnådd viser at hvis løver og gnuer levde alene, ville økningen i antall byttedyr føre til en rask akselerasjon av deres predasjon av rovdyr. På grunn av gruppens levemåte skjer ikke dette, aktiviteten til rovdyr øker relativt sakte, og det totale predasjonsnivået forblir lavt. Ifølge forfatterne, støttet av en rekke indirekte bevis, er antallet ofre i Serengeti ikke begrenset av løver i det hele tatt, men av matressurser.
Hvis fordelene med kollektivisme for ofrene er ganske åpenbare, forblir spørsmålet åpent i forhold til løvene. Denne studien viste tydelig at gruppelivsstilen for et rovdyr har en alvorlig ulempe - faktisk på grunn av den får hver enkelt løve mindre byttedyr. Åpenbart bør denne ulempen kompenseres av noen svært betydelige fordeler. Tradisjonelt ble det antatt at den sosiale livsstilen til løver er forbundet med jakt på store dyr, som er vanskelige å takle selv med en løve alene. Nylig begynte imidlertid mange eksperter (inkludert forfatterne av artikkelen under diskusjon) å tvile på riktigheten av denne forklaringen. Etter deres mening er kollektiv handling bare nødvendig for løver når de jakter på bøfler, og løver foretrekker å håndtere andre typer byttedyr alene.
Mer plausibel er antakelsen om at stolthet er nødvendig for å regulere rene interne problemer, som er mange i en løves liv. For eksempel er barnedrap vanlig blant dem - drap av andres unger av hanner. Det er lettere for kvinner som holdes i en gruppe å beskytte barna sine mot overgripere. I tillegg er det mye lettere for en stolthet enn for en ensom løve å forsvare sitt jaktområde mot nabostolthet.
Kilde: John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. Gruppedannelse stabiliserer rovdyr-byttedyr-dynamikken // Natur. 2007. V. 449. S. 1041–1043.
Simulering systemer "Rovdyret-Offer"
Sammendrag >> Økonomisk og matematisk modellering... systemer « Rovdyret-Offer" Laget av Gizyatullin R.R gr.MP-30 Sjekket av Lisovets Yu.P MOSKVA 2007 Introduksjon Interaksjon... modell interaksjoner rovdyr og ofre på overflaten. Forenkling av antakelser. La oss prøve å sammenligne offer og rovdyret noen...
Rovdyret-Offer
Abstrakt >> ØkologiAnvendelser av matematisk økologi er system rovdyret-offer. Den sykliske oppførselen til dette systemer i et stasjonært miljø ble ... ved å introdusere en ekstra ikke-lineær interaksjoner mellom rovdyret og et offer. Den resulterende modellen har på...
Synopsis økologi
Abstrakt >> Økologifaktor for ofre. Derfor interaksjon « rovdyret–offer" er periodisk og er system Lotkas ligninger... skiftet er mye mindre enn i system « rovdyret–offer". Lignende interaksjoner er også observert i batsisk mimikk. ...
Interaksjonsmodeller av to slag
Hypoteser om Volterra. Analogier med kjemisk kinetikk. Volterra-modeller for interaksjoner. Klassifisering av typer interaksjoner Konkurranse. Rovdyr-byttedyr. Generaliserte artsinteraksjonsmodeller . Kolmogorov modell. MacArthurs modell for interaksjon mellom to arter av insekter. Parametrisk og faseportretter av Bazykin-systemet.
Den italienske matematikeren Vito Volterra regnes med rette som grunnleggeren av den moderne matematiske teorien om populasjoner, som utviklet den matematiske teorien om biologiske samfunn, hvis apparat er differensial- og integro-differensialligninger.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris, 1931). I de påfølgende tiårene utviklet befolkningsdynamikken seg hovedsakelig i tråd med ideene uttrykt i denne boken. Den russiske oversettelsen av Volterras bok ble utgitt i 1976 under tittelen "Mathematical Theory of the Struggle for Existence" med et etterord av Yu.M. Svirezhev, som diskuterer historien til utviklingen av matematisk økologi i perioden 1931-1976.
Volterras bok er skrevet slik bøker om matematikk er skrevet. Den formulerer først noen antakelser om de matematiske objektene som skal studeres, og deretter gjennomføres en matematisk studie av egenskapene til disse objektene.
Systemene studert av Volterra består av to eller flere typer. I noen tilfeller vurderes beholdningen av mat som brukes. Ligningene som beskriver interaksjonen mellom disse artene er basert på følgende representasjoner.
Hypoteser om Volterra
1. Mat er enten tilgjengelig i ubegrensede mengder, eller forsyningen over tid er strengt regulert.
2. Individer av hver art dør på en slik måte at en konstant andel eksisterende individer omkommer per tidsenhet.
3. Rovdyr spiser byttedyr, og i løpet av en tidsenhet er antall spiste byttedyr alltid proporsjonalt med sannsynligheten for å møte individer av disse to artene, d.v.s. produktet av antall rovdyr og antall byttedyr.
4. Hvis det er mat i begrensede mengder og flere arter som er i stand til å konsumere den, så er andelen mat som konsumeres av en art per tidsenhet proporsjonal med antall individer av denne arten, tatt med en viss koeffisient avhengig av arten (modeller av interspesifikk konkurranse).
5. Dersom en art lever av mat som er tilgjengelig i ubegrensede mengder, er økningen i antall arter per tidsenhet proporsjonal med antallet arter.
6. Hvis en art lever av mat som er tilgjengelig i begrensede mengder, så reguleres reproduksjonen av matforbruket, dvs. per tidsenhet er økningen proporsjonal med mengden mat som spises.
Analogier med kjemisk kinetikk
Disse hypotesene har nære paralleller med kjemisk kinetikk. I ligningene for populasjonsdynamikk, som i ligningene for kjemisk kinetikk, brukes "kollisjonsprinsippet" når reaksjonshastigheten er proporsjonal med produktet av konsentrasjonene av de reagerende komponentene.
Faktisk, ifølge hypotesene til Volterra, hastigheten prosess utryddelsen av hver art er proporsjonal med artens overflod. I kjemisk kinetikk tilsvarer dette en monomolekylær dekomponeringsreaksjon av et eller annet stoff, og i en matematisk modell til negative lineære termer på høyre side av ligningene.
I henhold til konseptene for kjemisk kinetikk er hastigheten på en bimolekylær reaksjon av interaksjonen mellom to stoffer proporsjonal med sannsynligheten for en kollisjon av disse stoffene, dvs. produktet av deres konsentrasjon. På samme måte, ifølge hypotesene til Volterra, er reproduksjonshastigheten for rovdyr (rovdyrs død) proporsjonal med sannsynligheten for møter mellom rovdyr og byttedyr, dvs. produktet av deres tall. I begge tilfeller vises bilineære termer i modellsystemet på høyre side av de tilsvarende ligningene.
Til slutt tilsvarer de lineære positive leddene på høyresiden av Volterra-ligningene, som tilsvarer befolkningsvekst under ubegrensede forhold, de autokatalytiske termene for kjemiske reaksjoner. En slik likhet av ligninger i kjemiske og økologiske modeller gjør det mulig å anvende de samme forskningsmetodene for matematisk modellering av populasjonskinetikk som for systemer av kjemiske reaksjoner.
Klassifisering av typer interaksjoner
I samsvar med hypotesene til Volterra, samspillet mellom to arter, hvorav antallet x 1 og x 2 kan beskrives med ligningene:
(9.1)
Her parametere en Jeg - veksthastighetskonstanter for arter, c Jeg- populasjons selvbegrensende konstanter (intraspesifikk konkurranse), b ij- artsinteraksjonskonstanter, (Jeg, j= 1,2). Tegnene til disse koeffisientene bestemmer typen interaksjon.
I den biologiske litteraturen er interaksjoner vanligvis klassifisert i henhold til mekanismene som er involvert. Mangfoldet her er enormt: ulike trofiske interaksjoner, kjemiske interaksjoner som eksisterer mellom bakterier og planktonalger, interaksjoner av sopp med andre organismer, rekkefølger av planteorganismer assosiert spesielt med konkurranse om sollys og med utviklingen av jord, etc. En slik klassifisering virker udefinerbar.
E . Odum, med tanke på modellene foreslått av V. Volterra, foreslo en klassifisering ikke etter mekanismer, men etter resultater. I henhold til denne klassifiseringen skal forhold vurderes som positive, negative eller nøytrale, avhengig av om forekomsten av en art øker, avtar eller forblir uendret i nærvær av en annen art. Deretter kan hovedtypene av interaksjoner presenteres i form av en tabell.
TYPER AV ARTS INTERAKSJON
|
SYMBIOSE |
b 12 ,b 21 >0 |
||
|
KOMMENSALISME |
b 12 ,>0, b 21 =0 |
||
|
ROVDYR-Byttedyr |
b 12 ,>0, b 21 <0 |
||
|
AMENSALISME |
b 12 ,=0, b 21 <0 |
||
|
KONKURRANSE |
b 12 , b 21 <0 |
||
|
NØYTRALISME |
b 12 , b 21 =0 |
Den siste kolonnen viser fortegnene til interaksjonskoeffisientene fra systemet (9.1)
Vurder hovedtypene av interaksjoner
KONKURRANSELIGNINGER:
Som vi så i forelesning 6, er konkurranseligningene:
 (9.2)
(9.2)
Stasjonære systemløsninger:
(1).
![]()
Opprinnelsen til koordinatene, for alle parametere i systemet, er en ustabil node.
(2).
![]() (9.3)
(9.3)
C den stasjonære tilstanden (9.3) er en sal kl en 1 >b 12 /Med 2 og
stabil knute kl en 1 12 /s 2 . Denne tilstanden betyr at arten dør ut hvis dens egen veksthastighet er mindre enn en kritisk verdi.
(3).
![]() (9.4)
(9.4)
C stasjonær løsning (9.4)¾ sal kl en 2 >b 21 /c 1 og en stabil knute kl en 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Den stasjonære tilstanden (9.5) karakteriserer sameksistensen av to konkurrerende arter og er en stabil node hvis relasjonen er oppfylt:
![]()
Dette innebærer ulikhet:
b 12
b 21
som lar oss formulere betingelsen for sameksistens av arter:
Produktet av interpopuer mindre enn produktet av koeffisienter innenfor populasjonsinteraksjon.
Faktisk, la de naturlige vekstratene til de to betraktede arteneen 1 , a 2 er det samme. Da er den nødvendige betingelsen for stabilitet
c 2 > b 12 ,c 1 >b 21 .
Disse ulikhetene viser at økningen i antallet til en av konkurrentene undertrykker dens egen vekst sterkere enn veksten til en annen konkurrent. Hvis forekomsten av begge arter er begrenset, delvis eller fullstendig, av forskjellige ressurser, er de ovennevnte ulikhetene gyldige. Hvis begge artene har nøyaktig samme behov, vil en av dem være mer levedyktig og fortrenge sin konkurrent.
Oppførselen til fasebanene til systemet gir en visuell representasjon av mulige utfall av konkurranse. Vi likestiller høyresiden av likningene til systemet (9.2) til null:
x 1 (en 1-c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (en 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
I dette tilfellet får vi ligninger for hovedisoklinene til systemet
x 2 = – b 21 x 1 / c 2 +en 2/c2, x 2 = 0
er ligningene til isokliner av vertikale tangenter.
x 2 = – c 1 x 1 /b12+ en 1 /b 12 , x 1 = 0
er ligningene til isokliner av vertikale tangenter. Punktene med parvise skjæringspunkter for isoklinene til vertikale og horisontale tangentsystemer er stasjonære løsninger av ligningssystemet (9.2.), og deres koordinater ![]() er stasjonære antall konkurrerende arter.
er stasjonære antall konkurrerende arter.
Den mulige plasseringen av hovedisoklinene i systemet (9.2) er vist i fig. 9.1. Ris. 9.1entilsvarer artens overlevelsex 1, fig. 9.1 b- artens overlevelsex 2, fig. 9.1 i– sameksistens av arter under tilstand (9.6). Figur 9.1Gdemonstrerer triggersystemet. Her avhenger utfallet av konkurransen av startforholdene. Den stasjonære tilstanden (9,5), som ikke er null for begge typer, er ustabil. Dette er salen som separatrixen passerer gjennom, og skiller overlevelsesområdene til hver av artene.

Ris. 9.1.Plasseringen av de viktigste isoklinene i faseportrettet av Volterra-konkurransesystemet av to typer (9.2) med forskjellige forhold mellom parametere. Forklaringer i teksten.
For å studere konkurransen mellom arter ble det utført eksperimenter på en rekke organismer. Vanligvis velges to nært beslektede arter og dyrkes sammen og hver for seg under strengt kontrollerte forhold. Med visse intervaller gjennomføres en fullstendig eller selektiv folketelling. Registrer data fra flere gjentatte eksperimenter og analyser. Studiene ble utført på protozoer (spesielt ciliater), mange arter av biller av slekten Tribolium, Drosophila og ferskvannskrepsdyr (daphnia). Mange eksperimenter er utført på mikrobielle populasjoner (se forelesning 11). Eksperimenter ble også utført i naturen, inkludert på planarer (Reynolds), to arter av maur (Pontin) og andre. 9.2. vekstkurvene for kiselalger som bruker samme ressurs (opptar samme økologiske nisje) vises. Når den dyrkes i monokultur Asterionella formosa når et konstant tetthetsnivå og opprettholder konsentrasjonen av ressursen (silikat) på et konstant lavt nivå. B. Når den dyrkes i monokultur Synedruina oppfører seg på lignende måte og holder silikatkonsentrasjonen på et enda lavere nivå. B. Med samdyrking (i duplikat) Synedruina utkonkurrerer Asterionella formosa. Angivelig Synedra

Ris. 9.2.Konkurranse i kiselalger. en - når den dyrkes i monokultur Asterionella formosa når et konstant tetthetsnivå og holder konsentrasjonen av ressursen (silikat) på et konstant lavt nivå. b - når den dyrkes i monokultur Synedruina oppfører seg på lignende måte og holder silikatkonsentrasjonen på et enda lavere nivå. i - i samdyrking (i duplikat) Synedruina utkonkurrerer Asterionella formosa. Angivelig Synedra vinner konkurransen på grunn av dens evne til å utnytte underlaget mer fullstendig (se også forelesning 11).
G. Gauses eksperimenter på studiet av konkurranse er viden kjent, og demonstrerer overlevelsen til en av de konkurrerende artene og lar ham formulere "loven om konkurranseutstenging". Loven sier at kun én art kan eksistere i én økologisk nisje. På fig. 9.3. resultatene av Gauses forsøk for to Parametium-arter som okkuperer samme økologiske nisje (fig. 9.3 a, b) og arter som okkuperer forskjellige økologiske nisjer (fig. 9.3. c) presenteres.

Ris. 9.3. en- Populasjonsvekstkurver for to arter Parametium i enkeltartskulturer. Svarte sirkler - P Aurelia, hvite sirkler - P. Caudatum
b- P aurelia og P vekstkurver. Caudatum i en blandet kultur.
Av Gause, 1934
Konkurransemodellen (9.2) har mangler, spesielt følger det at sameksistens av to arter kun er mulig dersom deres forekomst begrenses av ulike faktorer, men modellen angir ikke hvor store forskjellene må være for å sikre langsiktig sameksistens . Samtidig er det kjent at langvarig sameksistens i et miljø i endring krever en forskjell som når en viss verdi. Innføringen av stokastiske elementer i modellen (for eksempel innføringen av en ressursbruksfunksjon) gjør at vi kan studere disse problemstillingene kvantitativt.
Predator+bytte-system
 (9.7)
(9.7)
Her, i motsetning til (9.2), skiltene b 12 og b 21 - annerledes. Som i tilfellet med konkurranse, opprinnelsen
![]() (9.8)
(9.8)
er et enkelt punkt av typen ustabil knute. Tre andre mulige stasjonære tilstander:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Dermed er bare byttet (9.10), kun rovdyret (9.9) (hvis det har andre matkilder) og sameksistens av begge artene (9.11) mulig. Det siste alternativet har vi allerede vurdert i forelesning 5. Mulige typer faseportretter for rovdyr-bytte-systemet er vist i fig. 9.4.
Isoklinene til de horisontale tangentene er rette linjer
x 2 = – b 21 X 1 /c 2 + en 1/c2, X 2 = 0,
og isoklinene til de vertikale tangentene- rett
x 2 = - c 1 X 1 /b 12 + en 2 /b 12 , X 1 = 0.
De stasjonære punktene ligger i skjæringspunktet mellom isoklinene til de vertikale og horisontale tangentene.

Fra fig. 9.4 følgende er sett. rovdyr-bytte-system (9.7) kan ha en stabil likevektsposisjon, hvori o rom offerbefolkningen er fullstendig utryddet ( ) og bare rovdyr var igjen (punkt 2 i fig. 9.4 en). Åpenbart kan en slik situasjon bare realiseres hvis, i tillegg til typen ofre som vurderes, X 1 rovdyr X 2 - har ekstra strømkilder. Dette faktum gjenspeiles i modellen av det positive leddet på høyre side av ligningen for x 2 . Enkeltstående poeng(1) og (3) (fig. 9.4 en) er ustabile. Andre mulighet – en stabil stasjonær tilstand der rovdyrbestanden har dødd helt ut og bare ofre er igjen – stabilt punkt(3) (Fig. 9.4 6 ). Her er et spesielt poeng (1) – også en ustabil node.
Til slutt den tredje muligheten – stabil sameksistens av rovdyr- og byttedyrpopulasjoner (fig. 9.4 i), hvis stasjonære overflod er uttrykt av formlene (9.11).
Som for en enkelt populasjon (se forelesning 3), for modellen (9.7) det er mulig å utvikle en stokastisk modell, men den kan ikke løses eksplisitt. Derfor begrenser vi oss til generelle betraktninger. Anta for eksempel at likevektspunktet er i en viss avstand fra hver av aksene. Deretter for fasebaner som verdienex 1 , x 2 forbli tilstrekkelig stor, vil en deterministisk modell være ganske tilfredsstillende. Men hvis en eller annen variabel på et tidspunkt i fasebanen ikke er veldig stor, kan tilfeldige fluktuasjoner bli betydelige. De fører til at det representative punktet vil flytte til en av aksene, noe som betyr utryddelse av den tilsvarende arten.
Dermed viser den stokastiske modellen seg å være ustabil, siden den stokastiske "driften" før eller senere fører til utryddelse av en av artene. I denne typen modell dør rovdyret til slutt ut, enten ved en tilfeldighet eller fordi byttedyrbestanden blir eliminert først. Den stokastiske modellen av rovdyr-bytte-systemet forklarer Gauses eksperimenter godt (Gause, 1934), i hvilke ciliater Paramettum candatum fungerte som et bytte for et annet ciliat Didinium nasatum – rovdyret. Forventet i henhold til deterministiske ligninger (9.7) likevektstallene i disse forsøkene var omtrent bare fem individer av hver art, så det er ikke noe overraskende i det faktum at i hvert gjentatte forsøk døde enten rovdyr eller byttedyr (og deretter rovdyr) ut ganske raskt. Resultatene av forsøkene presenteres i fig. 9.5.

Ris. 9.5. Vekst Parametium caudatum og rov ciliater Dadinium nasutum. Fra : Gause G.F. Kampen for tilværelsen. Baltimore, 1934
Så, analysen av Volterra-modellene for artsinteraksjon viser at til tross for den store variasjonen av typer oppførsel av slike systemer, kan det ikke være noen udempede populasjonssvingninger i modellen til konkurrerende arter i det hele tatt. Imidlertid observeres slike svingninger i naturen og i eksperimentet. Behovet for deres teoretiske forklaring var en av grunnene til å formulere modellbeskrivelser i en mer generell form.
Generaliserte modeller for interaksjon av to typer
Det er foreslått et stort antall modeller som beskriver samspillet mellom arter, hvis høyre side av likningene var funksjoner av størrelsen på de interagerende populasjonene. Spørsmålet om å utvikle generelle kriterier for å bestemme hvilken type funksjoner som kan beskrive oppførselen til den midlertidige populasjonsstørrelsen, inkludert stabile fluktuasjoner, ble diskutert. De mest kjente av disse modellene er de til Kolmogorov (1935, revidert 1972) og Rosenzweig (1963).
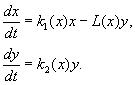 (9.12)
(9.12)
Modellen er basert på følgende forutsetninger:
1) Rovdyr samhandler ikke med hverandre, dvs. rovdyravlshastighet k 2 og antall ofre L, utryddet per tidsenhet av ett rovdyr, er ikke avhengig av y.
2) Økningen i antall byttedyr i nærvær av rovdyr er lik økningen i fravær av rovdyr minus antall byttedyr som er utryddet av rovdyr. Funksjoner k 1 (x), k 2 (x), L(x), er kontinuerlige og definert på den positive halvaksen x, y³ 0.
3) dk 1 /dx< 0. Dette betyr at multiplikasjonsfaktoren for byttedyr i fravær av et rovdyr avtar monotont med en økning i antall byttedyr, noe som gjenspeiler begrenset mat og andre ressurser.
4) dk 2 /dx> 0, k 2 (0) < 0 < k 2 (¥ ). Med en økning i antall byttedyr, avtar multiplikasjonsfaktoren for rovdyr monotont med en økning i antall byttedyr, og går fra negative verdier (når det ikke er noe å spise) til positive.
5) Antall ofre som er utryddet av ett rovdyr per tidsenhet L(x)> 0 på N> 0; L(0)=0.
Mulige typer faseportretter av systemet (9.12) er vist i fig. 9,6:

Ris. 9.6.Faseportretter av Kolmogorov-systemet (9.12), som beskriver samspillet mellom to typer for forskjellige parameterforhold. Forklaringer i teksten.
Stasjonære løsninger (det er to eller tre av dem) har følgende koordinater:
(1). ` x=0;` y=0.
Opprinnelsen til koordinatene for alle verdier av parameterne er en sadel (fig. 9.6 a-d).
(2). ` x=A,` y=0.(9.13)
ENbestemmes ut fra ligningen:
k 1 (EN)=0.
Stasjonær løsning (9.13) er en sal hvis B< EN (Fig. 9.6 en, b, G), B bestemt ut fra ligningen
k 2 (B)=0
Punkt (9.13) plasseres i den positive kvadranten if B>A . Dette er en stabil knute .
Det siste tilfellet, som tilsvarer rovdyrets død og byttets overlevelse, er vist i fig. 9.6 i.
(3). ` x=B,` y=C.(9.14)
Verdien av C bestemmes fra ligningene:
Punkt (9.14) - fokus (fig. 9.6 en) eller knute (fig. 9.6 G), hvis stabilitet avhenger av fortegnet for mengdens
s 2 = – k 1 (B)-k 1 (B)B+L(B)C.
Hvis en s>0, punktet er stabilt hviss<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
I utenlandsk litteratur blir en lignende modell foreslått av Rosenzweig og MacArthur (1963) oftere vurdert:
 (9.15)
(9.15)
hvor f(x) - endringshastigheten i antall ofre x i fravær av rovdyr, F( x,y) er intensiteten av predasjon, k- koeffisient som karakteriserer effektiviteten av byttedyrbiomassekonvertering til rovdyrbiomasse, e- Rovdyrdødelighet.
Modell (9.15) reduserer til et spesielt tilfelle av Kolmogorovs modell (9.12) under følgende forutsetninger:
1) antall rovdyr begrenses bare av antall byttedyr,
2) hastigheten som et gitt individ av et rovdyr spiser et bytte med, avhenger bare av rovdyrbestandstettheten og avhenger ikke av rovdyrbestandstettheten.
Så tar ligningene (9.15) formen.

Når man skal beskrive samspillet mellom virkelige arter, blir de riktige delene av ligningene konkretisert i samsvar med ideer om biologiske realiteter. Vurder en av de mest populære modellene av denne typen.
Modell for interaksjon mellom to arter av insekter (MacArthur, 1971)
Modellen, som vi skal diskutere nedenfor, ble brukt til å løse det praktiske problemet med skadedyrbekjempelse ved å sterilisere hanner av en av artene. Basert på de biologiske egenskapene til interaksjonen mellom arter, ble følgende modell skrevet
 (9.16)
(9.16)
Her x,y- biomasse av to arter av insekter. De trofiske interaksjonene til artene beskrevet i denne modellen er svært komplekse. Dette bestemmer formen til polynomene på høyre side av ligningene.
Tenk på høyre side av den første ligningen. Insektarter X spise larvene til arten på(medlem + k 3 y), men voksne av arten på spise larvene til arten X underlagt et stort antall arter X eller på eller begge typer (medlemmer – k 4 xy, – y 2). I det små X artsdødelighet X høyere enn dens naturlige økning (1 –k 1 +k 2 x–x 2 < 0 i det små X). I den andre ligningen, begrepet k 5 reflekterer artens naturlige vekst y; –k 6 y- selvbeherskelse av denne typen,–k 7 x- spise larver av arten på insekter av arten x, k 8 xy – vekst av arters biomasse på ved å bli spist av voksne insekter av arten på larver av arten X.
På fig. 9.7 grensesyklusen presenteres, som er banen til en stabil periodisk løsning av systemet (9.16).

Løsningen av spørsmålet om hvordan man sikrer sameksistensen av en populasjon med dets biologiske miljø, kan selvfølgelig ikke oppnås uten å ta hensyn til spesifikasjonene til et bestemt biologisk system og en analyse av alle dets sammenhenger. Samtidig gjør studiet av formelle matematiske modeller det mulig å svare på noen generelle spørsmål. Det kan hevdes at for modeller av typen (9.12) avhenger ikke populasjonenes kompatibilitet eller inkompatibilitet av deres opprinnelige størrelse, men bestemmes kun av arten av interaksjonen mellom arter. Modellen bidrar til å svare på spørsmålet: hvordan påvirke biocenosen, håndtere den for å ødelegge den skadelige arten så raskt som mulig.
Ledelsen kan reduseres til en kortsiktig, krampaktig endring i befolkningens størrelse X og y. Denne metoden tilsvarer metoder for kontroll som en enkelt ødeleggelse av en eller begge populasjoner med kjemiske midler. Fra uttalelsen formulert ovenfor, kan det sees at for kompatible populasjoner vil denne kontrollmetoden være ineffektiv, siden systemet over tid igjen vil nå et stasjonært regime.
En annen måte er å endre typen interaksjonsfunksjoner mellom typer, for eksempel når du endrer verdiene til systemparametere. Det er nettopp denne parametriske metoden som biologiske kampmetoder tilsvarer. Når steriliserte hanner blir introdusert, reduseres således koeffisienten for naturlig befolkningsvekst. Hvis vi samtidig får en annen type faseportrett, et hvor det kun er en stabil stasjonær tilstand med null skadedyrtall, vil kontrollen føre til ønsket resultat – ødeleggelse av skadedyrbestanden. Det er interessant å merke seg at noen ganger er det tilrådelig å bruke virkningen ikke på skadedyret selv, men på partneren. Hvilken av metodene som er mer effektiv, i det generelle tilfellet, er det umulig å si. Det avhenger av kontrollene som er tilgjengelige og av den eksplisitte formen til funksjonene som beskriver interaksjonen mellom populasjoner.
Modell A.D.Bazykin
Den teoretiske analysen av artsinteraksjonsmodeller er mest uttømmende utført i boken av A.D. Bazykin "Biophysics of interacting populations" (M., Nauka, 1985).
Tenk på en av rovdyr-bytte-modellene som er studert i denne boken.
 (9.17)
(9.17)
System (9.17) er en generalisering av den enkleste Volterra rovdyr-byttedyr-modellen (5.17) som tar hensyn til metningseffekten til rovdyr. Modell (5.17) antar at intensiteten av byttedyrbeite øker lineært med økende byttedyrtetthet, noe som ikke samsvarer med virkeligheten ved høye byttedyrtettheter. Ulike funksjoner kan velges for å beskrive rovdyrdiettens avhengighet av byttedyrtetthet. Det er viktigst at den valgte funksjonen med økende x tenderer asymptotisk til en konstant verdi. Modell (9.6) brukte logistisk avhengighet. I Bazykin-modellen er hyperbelen valgt som en slik funksjon x/(1+px). Husk at Monods formel, som beskriver avhengigheten av veksthastigheten til mikroorganismer på konsentrasjonen av substratet, har denne formen. Her fungerer byttet som et substrat, og rovdyret fungerer som mikroorganismer. .
System (9.17) avhenger av syv parametere. Antall parametere kan reduseres ved å endre variabler:
x® (A/D)x; y ® (A/D)/y;
t® (1/A)t; g (9,18)
og avhenger av fire parametere.
For en komplett kvalitativ studie er det nødvendig å dele det firdimensjonale parameterrommet inn i regioner med ulike typer dynamisk atferd, dvs. konstruere et parametrisk eller strukturelt portrett av systemet.
Deretter er det nødvendig å bygge faseportretter for hver av regionene i det parametriske portrettet og beskrive bifurkasjonene som oppstår med faseportretter ved grensene til ulike regioner i det parametriske portrettet.
Konstruksjonen av et komplett parametrisk portrett utføres i form av et sett med "skiver" (projeksjoner) av et parametrisk portrett av liten dimensjon med faste verdier for noen av parametrene.
Parametrisk portrett av systemet (9.18) for fast g og liten e vist i figur 9.8. Portrettet inneholder 10 områder med ulike typer fasebaneatferd.

Ris. 9.8.Parametrisk portrett av systemet (9.18) for fastg
og liten e
Oppførselen til systemet med forskjellige forhold mellom parametere kan være betydelig forskjellig (fig. 9.9). Følgende er mulig i systemet:
1) en stabil likevekt (region 1 og 5);
2) én stabil grensesyklus (region 3 og 8);
3) to stabile likevekter (region 2)
4) stabil grensesyklus og ustabil likevekt inne i den (region 6, 7, 9, 10)
5) stabil grensesyklus og stabil likevekt utenfor den (region 4).
I de parametriske områdene 7, 9, 10 er området med likevektsattraksjon begrenset av en ustabil grensesyklus som ligger inne i den stabile. Det mest interessante er faseportrettet som tilsvarer region 6 i det parametriske portrettet. Det er vist i detalj i fig. 9.10.

Tiltrekningsområdet for likevekt B 2 (skyggelagt) er en "snegl" som vrider seg fra det ustabile fokuset B 1 . Hvis det er kjent at systemet i det første øyeblikket var i nærheten av В 1, så er det mulig å bedømme om den tilsvarende banen vil komme til likevekt В 2 eller til en stabil grensesyklus som omgir de tre likevektspunktene С ( sal), В 1 og В 2 bare basert på sannsynlighetsbetraktninger.

Fig.9.10.Faseportrett av system 9.18 for parametrisk region 6. Attraksjonsområde B 2 er skyggelagt
På et parametrisk portrett(9.7) det er 22 ulike bifurkasjonsgrenser som dannes 7 forskjellige typer bifurkasjoner. Studien deres gjør det mulig å identifisere mulige typer systematferd når parametrene endres. For eksempel ved flytting fra området 1 til område 3 det er en fødsel av en liten grensesyklus, eller en myk fødsel av selvsvingninger rundt en enkelt likevekt PÅ. En lignende myk fødsel av selvsvingninger, men rundt en av likevektene, nemlig B 1 , oppstår når du krysser grensen til regioner 2 og 4. Ved flytting fra området 4 til område 5 stabil grensesyklus rundt et punktB 1 "bursts" på separatrix-løkken og det eneste tiltrekningspunktet er likevekten B 2 etc.
Av spesiell interesse for praksis er selvfølgelig utviklingen av kriterier for et systems nærhet til bifurkasjonsgrenser. Faktisk er biologer godt klar over egenskapen "buffer" eller "fleksibilitet" til naturlige økologiske systemer. Disse begrepene betegner vanligvis systemets evne til å absorbere ytre påvirkninger, så å si. Så lenge intensiteten av den ytre handlingen ikke overstiger en viss kritisk verdi, gjennomgår ikke systemets oppførsel kvalitative endringer. På faseplanet tilsvarer dette systemets retur til en stabil likevektstilstand eller til en stabil grensesyklus, hvis parametere ikke skiller seg mye fra den opprinnelige. Når intensiteten av påvirkningen overstiger den tillatte, "bryter systemet sammen", går over i en kvalitativt annen modus for dynamisk oppførsel, for eksempel dør det ganske enkelt ut. Dette fenomenet tilsvarer en bifurkasjonsovergang.
Hver type bifurkasjonsoverganger har sine egne særtrekk som gjør det mulig å bedømme faren for en slik overgang for økosystemet. Her er noen generelle kriterier som vitner om nærheten til en farlig grense. Som for én art, hvis en reduksjon i antallet av en av artene får systemet til å "sette seg fast" nær et ustabilt setepunkt, noe som uttrykkes i en veldig langsom gjenoppretting av antallet til den opprinnelige verdien, så systemet er nær den kritiske grensen. Endringen i form av svingninger i antall rovdyr og byttedyr fungerer også som en fareindikator. Hvis svingninger blir avslappende fra nær harmoniske, og amplituden til svingningene øker, kan dette føre til tap av stabilitet i systemet og utryddelse av en av artene.
Ytterligere utdyping av den matematiske teorien om samspillet mellom arter går langs linjen med å detaljere strukturen til selve bestandene og ta hensyn til tidsmessige og romlige faktorer.
Litteratur.
Kolmogorov A.N. Kvalitativ studie av matematiske modeller for populasjonsdynamikk. // Problemer med kybernetikk. M., 1972, utgave 5.
MacArtur R. Grafisk analyse av økologiske systemer// Biologisk avdelingsrapport Perinceton University. 1971
AD Bazykin "Biofysikk av interagerende populasjoner". M., Nauka, 1985.
W. Volterra: "Matematisk teori om kampen for tilværelsen." M.. Vitenskap, 1976
Gasbind G.F. Kampen for tilværelsen. Baltimore, 1934.
Her, i motsetning til (3.2.1), er tegnene (-012) og (+a2i) forskjellige. Som i tilfellet med konkurranse (ligningssystem (2.2.1)), er origo (1) for dette systemet et enkeltpunkt av typen "ustabil node". Tre andre mulige stasjonære tilstander:

Biologisk mening krever positive verdier X y x 2. For uttrykk (3.3.4) betyr dette at
Hvis koeffisienten for intraspesifikk konkurranse av rovdyr en,22 = 0, tilstand (3.3.5) fører til tilstanden ai2
Mulige typer faseportretter for ligningssystemet (3.3.1) er vist i fig. 3,2 a-c. Isoklinene til de horisontale tangentene er rette linjer
og isoklinene til de vertikale tangentene er rette
Fra fig. 3.2 viser følgende. Rovdyr-bytte-systemet (3.3.1) kan ha en stabil likevekt der byttedyrbestanden er fullstendig utryddet (x = 0) og kun rovdyr var igjen (punkt 2 i fig. 3.26). Åpenbart kan en slik situasjon bare realiseres hvis, i tillegg til typen ofre som vurderes, X rovdyret X2 har ekstra strømforsyninger. Dette faktum gjenspeiles i modellen av det positive leddet på høyre side av ligningen for xs. Enkeltpunkter (1) og (3) (fig. 3.26) er ustabile. Den andre muligheten er en stabil stasjonær tilstand, der bestanden av rovdyr døde helt ut og bare ofre ble igjen - et stabilt punkt (3) (fig. 3.2a). Her er singularpunktet (1) også en ustabil node.
Til slutt er den tredje muligheten stabil sameksistens av rovdyr- og byttedyrpopulasjoner (fig. 3.2 c), hvis stasjonære tall er uttrykt med formler (3.3.4). La oss vurdere denne saken mer detaljert.
Anta at koeffisientene for intraspesifikk konkurranse er lik null (ai= 0, i = 1, 2). La oss også anta at rovdyr bare lever av byttedyr av arten X og i deres fravær dør de ut med en hastighet på C2 (i (3.3.5) C2
La oss gjennomføre en detaljert studie av denne modellen ved å bruke notasjonen som er mest akseptert i litteraturen. Oppusset

Ris. 3.2. Plasseringen av de viktigste isoklinene i faseportrettet av Volterra-systemets rovdyr-bytte for forskjellige forhold mellom parametere: en- Om -
FRA Jeg C2 C2
1, 3 - ustabilt, 2 - stabilt entallspunkt; i -
1, 2, 3 - ustabilt, 4 - stabilt entallspunkt signifikant
Rovdyr-bytte-systemet i disse notasjonene har formen:

Vi skal studere egenskapene til løsninger til system (3.3.6) på faseplanet N1
PÅ2
Systemet har to stasjonære løsninger. De er enkle å bestemme ved å likestille høyresiden av systemet til null. Vi får: 
Derav de stasjonære løsningene:

La oss se nærmere på den andre løsningen. La oss finne det første integralet av systemet (3.3.6) som ikke inneholder t. Multipliser den første ligningen med -72, den andre med -71 og legg til resultatene. Vi får:
Nå deler vi den første ligningen med N og gange med € 2, og del den andre med JV 2 og gang med e. La oss legge til resultatene igjen:
Ved å sammenligne (3.3.7) og (3.3.8), vil vi ha:

Ved å integrere får vi:

Dette er den ønskede første integralen. Systemet (3.3.6) er altså konservativt, siden det har det første integralet av bevegelse, en mengde som er en funksjon av variablene i systemet N og N2 og uavhengig av tid. Denne egenskapen gjør det mulig å konstruere et konseptsystem for Volterra-systemer som ligner på statistisk mekanikk (se kapittel 5), hvor en vesentlig rolle spilles av størrelsen på energien til systemet, som er uendret i tid.
For hver faste c > 0 (som tilsvarer visse startdata), tilsvarer integralet en viss bane på planet N1 PÅ2 , tjener som banen til systemet (3.3.6).
Tenk på en grafisk metode for å konstruere en bane, foreslått av Volterra selv. Merk at høyre side av formelen (3.3.9) bare avhenger av D r 2, og venstre side avhenger kun av N. Betegn
Av (3.3.9) følger at mellom X og Y det er et forholdsmessig forhold
På fig. 3.3 viser de første kvadrantene av fire koordinatsystemer XOY, NEI, N2 OKSE og D G 1 0N2 slik at de alle har et felles opphav.
I øvre venstre hjørne (kvadrant NEI) grafen til funksjonen (3.3.8) er konstruert, nederst til høyre (kvadrant N2 okse)- funksjonsgraf Y. Den første funksjonen har min kl Ni = og den andre - maks kl N2 = ?-
Til slutt, i kvadranten XOY konstruer linjen (3.3.12) for noen faste FRA.
Merk et poeng N på aksel PÅ. Dette punktet tilsvarer en viss verdi J(N 1), som er lett å finne ved å tegne en vinkelrett

Ris. 3.3.
gjennom N til den skjærer kurven (3.3.10) (se fig. 3.3). I sin tur tilsvarer verdien av K(A^) et punkt M på linjen Y = cX og dermed en viss verdi X(N) = Y(N)/c som kan finnes ved å tegne perpendikulære ER og MD. Den funnet verdien (dette punktet er markert i figuren med bokstaven D) matche to poeng R og G på kurven (3.3.11). Ved å tegne perpendikulære punkter finner vi to punkter samtidig E" og E" liggende på kurven (3.3.9). Koordinatene deres er:

Tegning vinkelrett ER, har vi krysset kurven (3.3.10) på ett punkt til PÅ. Dette punktet tilsvarer det samme R og Q på kurven (3.3.11) og det samme N og SCH. Koordinere N dette punktet kan finnes ved å slippe vinkelrett fra PÅ per aksel PÅ. Så vi får poeng F" og F" som også ligger på kurven (3.3.9).
Kommer fra et annet punkt N, på samme måte får vi en ny firedobbel av punkter som ligger på kurven (3.3.9). Unntaket er prikken Ni= ?2/72- Basert på det får vi bare to poeng: Til og L. Dette vil være de nedre og øvre punktene på kurven (3.3.9).
Kan ikke komme fra verdier N, og fra verdiene N2 . På vei fra N2 til kurven (3.3.11), for så å stige til den rette linjen Y = cX, og derfra krysse kurven (3.3.10), finner vi også fire punkter på kurven (3.3.9). Unntaket er prikken Nei=?1/71- Basert på det får vi bare to poeng: G og TIL. Dette vil være punktene lengst til venstre og lengst til høyre på kurven (3.3.9). Ved å spørre annerledes N og N2 og etter å ha mottatt nok poeng, koble dem sammen, konstruerer vi omtrent kurven (3.3.9).
Det kan sees fra konstruksjonen at dette er en lukket kurve som inneholder punktet 12 = (?2/721? N yu og N20. Å ta en annen verdi av C, dvs. andre startdata, får vi en annen lukket kurve som ikke skjærer den første og som også inneholder punktet (?2/721?1/71)1 i seg selv. Dermed er familien av baner (3.3.9) familien av lukkede linjer som omgir punktet 12 (se fig. 3.3). Vi undersøker typen stabilitet til dette ene punktet ved å bruke Lyapunov-metoden.
Siden alle parametere e 1, ?2, 71,72 er positive, pr (N[ er plassert i den positive kvadranten av faseplanet. Linearisering av systemet nær dette punktet gir:

Her n(t) og 7i2(N1, N2 :
Karakteristisk ligning for systemet (3.3.13):

Røttene til denne ligningen er rent imaginære:

Studiet av systemet viser således at banene nær entallspunktet er representert av konsentriske ellipser, og selve entallspunktet er sentrum (fig. 3.4). Volterra-modellen som vurderes har også lukkede baner langt fra singularpunktet, selv om formen på disse banene allerede er forskjellig fra ellipsoidale. Variabel oppførsel Ni, N2 i tid er vist i fig. 3.5.

Ris. 3.4.

Ris. 3.5. Avhengigheten av antall byttedyr N jeg og rovdyr N2 fra tid
Et enkelt punkt av typen sentrum er stabilt, men ikke asymptotisk. La oss bruke dette eksemplet for å vise hva det er. La vibrasjonene Ni(t) og LGgM oppstår på en slik måte at det representative punktet beveger seg langs faseplanet langs bane 1 (se fig. 3.4). I det øyeblikket punktet er i posisjon M, legges et visst antall individer til systemet fra utsiden N 2 slik at det representative punktet hopper fra punktet M punkt A/". Etter det, hvis systemet igjen er overlatt til seg selv, svingene Ni og N2 vil allerede oppstå med større amplituder enn før, og det representative punktet beveger seg langs bane 2. Dette betyr at svingningene i systemet er ustabile: de endrer karakteristikker permanent under ytre påvirkning. I det følgende tar vi for oss modeller som beskriver stabile oscillerende regimer og viser at slike asymptotiske stabile periodiske bevegelser er representert på faseplanet ved hjelp av grensesykluser.
På fig. 3.6 viser eksperimentelle kurver - svingninger i antall pelsbærende dyr i Canada (ifølge Hudson's Bay Company). Disse kurvene er bygget på grunnlag av data om antall høstede skinn. Periodene med svingninger i antall harer (byttedyr) og gauper (rovdyr) er omtrent like og er i størrelsesorden 9-10 år. Samtidig er maksimalt antall harer som regel ett år foran maksimalt antall gauper.
Formen på disse eksperimentelle kurvene er mye mindre korrekte enn de teoretiske. Men i dette tilfellet er det tilstrekkelig at modellen sikrer sammenfall av de mest betydningsfulle egenskapene til de teoretiske og eksperimentelle kurvene, dvs. amplitudeverdier og faseskift mellom fluktuasjoner i antall rovdyr og byttedyr. En mye mer alvorlig mangel ved Volterra-modellen er ustabiliteten til løsninger på ligningssystemet. Faktisk, som nevnt ovenfor, bør enhver tilfeldig endring i forekomsten av en eller annen art føre, etter modellen, til en endring i amplituden til svingningene til begge artene. Naturligvis, under naturlige forhold, blir dyr utsatt for et utallig antall slike tilfeldige påvirkninger. Som det fremgår av forsøkskurvene, varierer amplituden av fluktuasjoner i antall arter lite fra år til år.
Volterra-modellen er en referanse (grunnleggende) modell for matematisk økologi i samme grad som den harmoniske oscillatormodellen er grunnleggende for klassisk og kvantemekanikk. Ved hjelp av denne modellen, basert på svært forenklede ideer om arten av mønstrene som beskriver oppførselen til systemet, rent matematisk
kapittel 3

Ris. 3.6. Kinetiske kurver for overflod av pelsbærende dyr I følge Hudson's Bay Fur Company (Seton-Thomson, 1987) ble det trukket en konklusjon ved hjelp av kalkulus om den kvalitative karakteren av oppførselen til et slikt system - om tilstedeværelsen av populasjonssvingninger i et slikt system. Uten konstruksjonen av en matematisk modell og bruken av den, ville en slik konklusjon vært umulig.
I den enkleste formen vi har vurdert ovenfor, har Volterra-systemet to grunnleggende og sammenhengende mangler. Deres "eliminering" er viet omfattende økologisk og matematisk litteratur. For det første endrer inkluderingen i modellen av eventuelle, vilkårlig små, tilleggsfaktorer kvalitativt systemets oppførsel. Den andre "biologiske" ulempen med modellen er at den ikke inkluderer de grunnleggende egenskapene som er iboende i noen par av populasjoner som samhandler i henhold til rovdyr-byttedyr-prinsippet: effekten av metning av rovdyret, de begrensede ressursene til rovdyret og byttedyret. med overskudd av byttedyr, mulighet for et minimum antall byttedyr tilgjengelig for rovdyr osv.
For å eliminere disse ulempene har forskjellige modifikasjoner av Volterra-systemet blitt foreslått av forskjellige forfattere. De mest interessante av dem vil bli vurdert i avsnitt 3.5. Her dveler vi kun ved en modell som tar hensyn til selvbegrensninger i veksten til begge populasjoner. Eksemplet med denne modellen viser tydelig hvordan løsningenes natur kan endres når systemparametrene endres.
Så vi vurderer systemet

System (3.3.15) skiller seg fra det tidligere vurderte systemet (3.3.6) ved tilstedeværelsen av ledd på formen -7 på høyre side av ligningene uNf,
Disse medlemmene gjenspeiler det faktum at rovdyrbestanden ikke kan vokse i det uendelige selv i fravær av rovdyr på grunn av begrensede matressurser, begrenset eksistensspekter. De samme "selvbegrensningene" pålegges bestanden av rovdyr.
For å finne de stasjonære antall arter iVi og N2 lik null de riktige delene av systemlikningene (3.3.15). Løsninger med null antall rovdyr eller byttedyr vil ikke interessere oss nå. Tenk derfor på et algebraisk system
ligninger  Hennes avgjørelse
Hennes avgjørelse
gir oss koordinatene til entallspunktet. Her bør tilstanden til positiviteten til stasjonære tall settes på parametrene til systemet: N> 0 og N2 > 0. Røttene til den karakteristiske ligningen til et system linearisert i et nabolag til et enkeltpunkt (3.3.16):

Det kan sees av uttrykket for de karakteristiske tallene at dersom tilstanden
da utfører antallet rovdyr og byttedyr dempet svingninger i tid, systemet har et enkeltpunkt som ikke er null og et stabilt fokus. Faseportrettet av et slikt system er vist i fig. 3,7 a.
La oss anta at parametrene i ulikhet (3.3.17) endrer sine verdier på en slik måte at tilstand (3.3.17) blir en likhet. Da er de karakteristiske tallene til systemet (3.3.15) like, og dets entallspunkt vil ligge på grensen mellom områdene med stabile foci og noder. Når ulikhetstegnet (3.3.17) reverseres, blir entallspunktet en stabil node. Faseportrettet av systemet for dette tilfellet er vist i fig. 3,76.
Som for en enkelt populasjon kan en stokastisk modell utvikles for modell (3.3.6), men den kan ikke løses eksplisitt. Derfor begrenser vi oss til generelle betraktninger. Anta for eksempel at likevektspunktet er i en viss avstand fra hver av aksene. Så for fasebaner der verdiene til JVj, N2 forbli tilstrekkelig stor, vil en deterministisk modell være ganske tilfredsstillende. Men hvis på et tidspunkt


Ris. 3.7. Faseportrett av systemet (3.3.15): en - når forholdet (3.3.17) mellom parameterne er oppfylt; b- når du utfører det omvendte forholdet mellom parameterne
fasebane, er enhver variabel ikke veldig stor, så kan tilfeldige svingninger bli betydelige. De fører til at det representative punktet vil flytte til en av aksene, noe som betyr utryddelse av den tilsvarende arten. Dermed viser den stokastiske modellen seg å være ustabil, siden den stokastiske "driften" før eller siden fører til utryddelse av en av artene. I denne typen modell dør rovdyret til slutt ut, enten ved en tilfeldighet eller fordi byttedyrbestanden blir eliminert først. Den stokastiske modellen av rovdyr-bytte-systemet forklarer godt forsøkene til Gause (Gause, 1934; 2000), der ciliater Paramettum candatum fungerte som et bytte for et annet ciliat Didinium nasatum- rovdyret. Likevektstallene forventet i henhold til deterministiske ligninger (3.3.6) i disse eksperimentene var omtrent bare fem individer av hver art, så det er ikke noe overraskende i det faktum at i hvert gjentatte forsøk døde enten rovdyr eller byttedyr ut ganske raskt (og deretter rovdyr).
Så, analysen av Volterra-modellene for artsinteraksjon viser at til tross for den store variasjonen av typer oppførsel av slike systemer, kan det ikke være noen udempede populasjonssvingninger i modellen til konkurrerende arter i det hele tatt. I rovdyr-bytte-modellen oppstår udempede svingninger på grunn av valg av en spesiell form for modellligningene (3.3.6). I dette tilfellet blir modellen ikke-grov, noe som indikerer fraværet av mekanismer i et slikt system som søker å bevare sin tilstand. Imidlertid observeres slike svingninger i naturen og eksperimentet. Behovet for deres teoretiske forklaring var en av grunnene til å formulere modellbeskrivelser i en mer generell form. Avsnitt 3.5 er viet vurdering av slike generaliserte modeller.