Mô hình toán học kẻ săn mồi bảo thủ. Bài học: Nghiên cứu định tính mô hình kẻ săn mồi - con mồi. Mô hình mô phỏng của hệ thống "Predator-Prey"
Động vật ăn thịt có thể ăn động vật ăn cỏ, và cả những động vật săn mồi yếu ớt. Động vật ăn thịt có nhiều loại thức ăn, dễ dàng chuyển từ con mồi này sang con mồi khác, dễ tiếp cận hơn. Những kẻ săn mồi thường tấn công những con mồi yếu ớt. Sự cân bằng sinh thái được duy trì giữa các quần thể động vật ăn thịt con mồi. […]
Nếu trạng thái cân bằng không ổn định (không có chu kỳ giới hạn) hoặc chu kỳ bên ngoài không ổn định, thì số lượng của cả hai loài, bị biến động mạnh, rời khỏi vùng lân cận của trạng thái cân bằng. Hơn nữa, sự thoái hóa nhanh chóng (trong tình huống đầu tiên) xảy ra với sự thích nghi thấp của động vật ăn thịt, tức là với tỷ lệ tử vong cao của nó (so với tốc độ sinh sản của nạn nhân). Điều này có nghĩa là một kẻ săn mồi yếu ớt về mọi mặt sẽ không góp phần vào sự ổn định của hệ thống và tự nó chết đi. […]
Áp lực của kẻ thù đặc biệt mạnh khi, trong đồng tiến hóa giữa kẻ săn mồi và con mồi, sự cân bằng dịch chuyển về phía kẻ săn mồi và phạm vi của con mồi thu hẹp. Cuộc đấu tranh có liên quan mật thiết đến việc thiếu nguồn thức ăn, nó cũng có thể là cuộc đấu tranh trực tiếp, ví dụ, của những kẻ săn mồi lấy không gian như một nguồn tài nguyên, nhưng thường thì nó chỉ đơn giản là sự di dời của một loài không có đủ thức ăn trong một lãnh thổ của một loài có đủ lượng thức ăn như nhau. Đây là cuộc thi giữa các loài giữa các loài. […]
 |
Cuối cùng, trong hệ thống “động vật ăn thịt-con mồi” được mô tả bởi mô hình (2.7), sự xuất hiện của sự không ổn định khuếch tán (với sự ổn định cân bằng cục bộ) chỉ có thể xảy ra nếu tỷ lệ tử vong tự nhiên của động vật ăn thịt tăng theo quần thể của nó nhanh hơn hàm tuyến tính, và chức năng dinh dưỡng khác với Volterra hoặc khi quần thể con mồi là quần thể kiểu Ollie. […]
Về mặt lý thuyết, trong mô hình "một kẻ săn mồi - hai con mồi", sự săn mồi tương đương (không ưa thích một hoặc một loại con mồi khác) có thể ảnh hưởng đến sự chung sống cạnh tranh của các loài săn mồi chỉ ở những nơi đã tồn tại trạng thái cân bằng ổn định tiềm tàng. Sự đa dạng chỉ có thể tăng lên trong những điều kiện mà các loài kém khả năng cạnh tranh có tốc độ gia tăng quần thể cao hơn các loài ưu thế. Điều này làm cho chúng ta có thể hiểu được tình hình khi ngay cả việc chăn thả cũng dẫn đến sự gia tăng tính đa dạng của các loài thực vật khi một số lượng lớn hơn các loài đã được lựa chọn để sinh sản nhanh chóng cùng tồn tại với các loài mà quá trình tiến hóa nhằm tăng khả năng cạnh tranh. [...]
Tương tự như vậy, việc lựa chọn con mồi, tùy thuộc vào mật độ của nó, có thể dẫn đến trạng thái cân bằng ổn định trong các mô hình lý thuyết về hai loại con mồi cạnh tranh, nơi không tồn tại trạng thái cân bằng trước đó. Để làm được điều này, động vật ăn thịt phải có khả năng đáp ứng chức năng và số lượng đối với những thay đổi về mật độ con mồi; Tuy nhiên, có thể là việc chuyển đổi (các cuộc tấn công thường xuyên không tương xứng vào nạn nhân nhiều nhất) sẽ quan trọng hơn trong trường hợp này. Thật vậy, chuyển mạch được phát hiện có tác dụng ổn định trong hệ thống "một kẻ săn mồi - n con mồi" và là cơ chế duy nhất có khả năng ổn định tương tác trong trường hợp các hốc con mồi hoàn toàn trùng nhau. Vai trò này có thể được thực hiện bởi những kẻ săn mồi không chuyên biệt. Sự ưa thích của những kẻ săn mồi chuyên biệt hơn đối với một đối thủ cạnh tranh có ưu thế hoạt động giống như sự chuyển đổi của động vật ăn thịt và có thể ổn định các tương tác lý thuyết trong các mô hình mà trước đó không có sự cân bằng giữa các loài săn mồi, miễn là các hốc của chúng được tách biệt ở một mức độ nào đó. [.. .]
Ngoài ra, cộng đồng không được ổn định và kẻ săn mồi 'mạnh về mọi mặt', tức là thích nghi tốt với một con mồi nhất định và tỷ lệ tử vong tương đối thấp. Trong trường hợp này, hệ thống có một chu kỳ giới hạn không ổn định và, mặc dù vị trí cân bằng ổn định, sẽ bị thoái hóa trong một môi trường ngẫu nhiên (động vật ăn thịt ăn thịt con mồi và kết quả là chết). Tình trạng này tương ứng với sự thoái hóa chậm. […]
Do đó, với sự thích nghi tốt của động vật ăn thịt trong vùng lân cận của trạng thái cân bằng ổn định, các chu kỳ không ổn định và ổn định có thể phát sinh, tức là tùy thuộc vào các điều kiện ban đầu, hệ thống “động vật ăn thịt-con mồi” hoặc có xu hướng cân bằng, hoặc dao động, rời khỏi nó, hoặc sự dao động ổn định về số lượng của cả hai loài được thiết lập trong vùng lân cận của trạng thái cân bằng. [...]
Các sinh vật được xếp vào nhóm săn mồi ăn các sinh vật khác, tiêu diệt con mồi của chúng. Do đó, giữa các sinh vật sống, cần phân biệt thêm một hệ thống phân loại, đó là “động vật ăn thịt” và “nạn nhân”. Mối quan hệ giữa các sinh vật như vậy đã phát triển trong suốt quá trình tiến hóa của sự sống trên hành tinh của chúng ta. Sinh vật ăn thịt đóng vai trò là cơ quan điều hòa tự nhiên về số lượng sinh vật bắt mồi. Sự gia tăng số lượng "động vật ăn thịt" dẫn đến giảm số lượng "con mồi", do đó, làm giảm nguồn cung cấp thức ăn ("con mồi") cho "động vật ăn thịt", điều này nói chung làm giảm số lượng của "con mồi", v.v ... Như vậy, trong sinh vật học có sự biến động liên tục về số lượng vật ăn thịt và con mồi, nói chung, một sự cân bằng nhất định được thiết lập trong một thời gian nhất định trong điều kiện môi trường khá ổn định. [... ]
Điều này cuối cùng dẫn đến sự cân bằng sinh thái giữa quần thể động vật ăn thịt và con mồi. […]
Đối với chức năng dinh dưỡng thuộc loại thứ ba, trạng thái cân bằng sẽ ổn định nếu N là điểm uốn của chức năng (xem Hình 2, c). Điều này xuất phát từ thực tế là chức năng dinh dưỡng bị lõm vào trong khoảng thời gian và do đó, tỷ lệ tiêu thụ con mồi tương đối của động vật ăn thịt tăng lên. [...]
Cho Гг = -Г, tức là có một cộng đồng thuộc kiểu “kẻ săn mồi”. Trong trường hợp này, số hạng đầu tiên trong biểu thức (7.4) bằng 0, và để đáp ứng điều kiện ổn định đối với xác suất của trạng thái cân bằng N, thì số hạng thứ hai cũng không phải là số dương. [ ...]
Do đó, đối với cộng đồng được coi là thuộc loại động vật ăn thịt-con mồi, chúng ta có thể kết luận rằng trạng thái cân bằng dương nói chung là ổn định tiệm cận, tức là, đối với bất kỳ dữ liệu ban đầu nào với điều kiện N> 0. [...]
Vì vậy, trong một môi trường đồng nhất không có nơi trú ẩn để sinh sản, loài săn mồi sớm muộn gì cũng tiêu diệt quần thể con mồi rồi tự chết. Sóng sự sống ”(sự thay đổi số lượng kẻ săn mồi và con mồi) nối tiếp nhau với sự thay đổi liên tục theo giai đoạn, và trung bình số lượng cả kẻ săn mồi và con mồi vẫn xấp xỉ ở cùng một mức. Thời gian của giai đoạn này phụ thuộc vào tốc độ tăng trưởng của cả hai loài và vào các thông số ban đầu. Đối với quần thể con mồi, ảnh hưởng của động vật ăn thịt là tích cực, vì sự sinh sản quá mức của nó sẽ dẫn đến sự suy giảm số lượng của chúng. Đổi lại, tất cả các cơ chế ngăn chặn việc tiêu diệt hoàn toàn con mồi đều góp phần vào việc bảo tồn cơ sở thức ăn của động vật ăn thịt. [...]
Các sửa đổi khác có thể là do hành vi của động vật ăn thịt. Số lượng cá thể con mồi mà một kẻ săn mồi có thể tiêu thụ tại một thời điểm nhất định có giới hạn của nó. Ảnh hưởng của sự bão hòa của động vật ăn thịt khi tiếp cận ranh giới này được thể hiện trong Bảng. 2-4, B. Tương tác được mô tả bởi phương trình 5 và 6 có thể có điểm cân bằng ổn định hoặc biểu hiện dao động tuần hoàn. Tuy nhiên, các chu kỳ như vậy khác với những chu kỳ được phản ánh trong các phương trình Lotka-Volterra 1 và 2. Các chu kỳ được chuyển tải bởi các phương trình 5 và 6 có thể có biên độ và mật độ trung bình không đổi miễn là môi trường không đổi; sau khi vi phạm xảy ra, chúng có thể trở lại biên độ và mật độ trung bình trước đó của chúng. Các chu kỳ như vậy, được phục hồi sau khi vi phạm, được gọi là chu kỳ giới hạn ổn định. Tương tác của thỏ rừng và linh miêu có thể được coi là một chu kỳ giới hạn ổn định, nhưng đây không phải là chu kỳ Lotka-Volterra. […]
Chúng ta hãy xem xét sự xuất hiện của sự mất ổn định khuếch tán trong hệ thống "kẻ săn mồi-con mồi", nhưng trước tiên chúng ta viết ra các điều kiện đảm bảo sự xuất hiện của sự mất ổn định khuếch tán trong hệ thống (1.1) tại n = 2. Rõ ràng là cân bằng (N , W) là cục bộ (tức là [...]
Chúng ta hãy chuyển sang việc giải thích các trường hợp liên quan đến sự chung sống lâu dài của kẻ săn mồi và con mồi. Rõ ràng là trong trường hợp không có chu kỳ giới hạn, trạng thái cân bằng ổn định sẽ tương ứng với sự biến động của quần thể trong một môi trường ngẫu nhiên, và biên độ của chúng sẽ tỷ lệ với sự phân tán của nhiễu loạn. Hiện tượng như vậy sẽ xảy ra nếu động vật ăn thịt có tỷ lệ tử vong tương đối cao, đồng thời mức độ thích nghi cao đối với con mồi nhất định. […]
Bây giờ chúng ta hãy xem xét động lực của hệ thống thay đổi như thế nào với sự gia tăng sức khỏe của động vật ăn thịt, tức là với việc giảm b từ 1 đến 0. Nếu thể lực đủ thấp, thì không có chu kỳ giới hạn và trạng thái cân bằng không ổn định. Với sự phát triển của thể lực trong vùng lân cận của trạng thái cân bằng này, sự xuất hiện của một chu kỳ ổn định và sau đó là một chu kỳ không ổn định bên ngoài là có thể xảy ra. Tùy thuộc vào các điều kiện ban đầu (tỷ lệ giữa sinh khối vật ăn thịt và con mồi), hệ thống có thể mất tính ổn định, tức là rời khỏi vùng lân cận của trạng thái cân bằng, hoặc các dao động ổn định sẽ được thiết lập trong nó theo thời gian. Sự phát triển hơn nữa về thể lực làm cho bản chất dao động của hành vi của hệ thống là không thể. Tuy nhiên, khi b […]
Một ví dụ về phản hồi tiêu cực (ổn định) là mối quan hệ giữa động vật ăn thịt và con mồi hoặc hoạt động của hệ thống cacbonat đại dương (dung dịch CO2 trong nước: CO2 + H2O -> H2CO3). Thông thường, lượng carbon dioxide hòa tan trong nước đại dương ở trạng thái cân bằng một phần với nồng độ carbon dioxide trong khí quyển. Sự gia tăng cục bộ của carbon dioxide trong khí quyển sau khi núi lửa phun trào dẫn đến việc tăng cường quang hợp và sự hấp thụ của nó bởi hệ thống cacbonat của đại dương. Khi mức độ carbon dioxide trong khí quyển giảm, hệ thống cacbonat của đại dương thải CO2 vào khí quyển. Do đó, nồng độ carbon dioxide trong khí quyển khá ổn định. […]
[ ...]
Như R. Ricklefs (1979) đã lưu ý, có những yếu tố góp phần vào sự ổn định của các mối quan hệ trong hệ thống “kẻ săn mồi - con mồi”: sự kém hiệu quả của kẻ săn mồi, sự hiện diện của các nguồn thức ăn thay thế trong động vật ăn thịt, giảm sự chậm trễ trong phản ứng của động vật ăn thịt, cũng như các hạn chế về môi trường do môi trường bên ngoài áp đặt lên một hoặc nhiều quần thể khác nhau. Mối quan hệ tương tác giữa quần thể vật ăn thịt và con mồi rất đa dạng và phức tạp. Do đó, nếu các động vật ăn thịt đủ hiệu quả, chúng có thể điều chỉnh mật độ của quần thể con mồi, giữ cho nó ở mức thấp hơn khả năng của môi trường. Thông qua ảnh hưởng của chúng đối với quần thể con mồi, động vật ăn thịt ảnh hưởng đến sự tiến hóa của các đặc điểm khác nhau của con mồi, cuối cùng dẫn đến sự cân bằng sinh thái giữa quần thể động vật ăn thịt và con mồi. [...]
Nếu một trong các điều kiện được đáp ứng: 0 1/2. Nếu 6> 1 (kA [...]
Sự ổn định của quần thể sinh vật và môi trường chỉ phụ thuộc vào sự tương tác giữa thực vật - sinh vật tự dưỡng và sinh vật dị dưỡng ăn cỏ. Động vật ăn thịt ở bất kỳ kích thước nào cũng không thể làm xáo trộn cân bằng sinh thái của quần xã, vì trong điều kiện tự nhiên, chúng không thể tăng số lượng với số lượng con mồi không đổi. Động vật ăn thịt không chỉ phải tự di chuyển mà chỉ có thể ăn những động vật đang di chuyển. […]
Không có loài cá nào khác phân bố rộng rãi như cá pike. Ở một vài khu vực đánh bắt ở những vùng nước đọng hoặc nước chảy, không có áp lực từ những con mồi để duy trì sự cân bằng giữa con mồi và kẻ săn mồi. Pike là đại diện đặc biệt tốt trên thế giới. Chúng được đánh bắt trên khắp bán cầu bắc) từ Hoa Kỳ và Canada ở Bắc Mỹ, qua châu Âu đến bắc Á. […]
Một khả năng khác của sự chung sống ổn định nảy sinh ở đây, trong một phạm vi hẹp của sự thích nghi tương đối cao. Khi chuyển sang một chế độ không ổn định với một động vật ăn thịt rất “tốt”, một chu kỳ giới hạn bên ngoài ổn định có thể phát sinh, trong đó sự tiêu tán sinh khối được cân bằng bởi dòng chảy của nó vào hệ thống (năng suất cao của con mồi). Sau đó, một tình huống tò mò nảy sinh khi có khả năng xảy ra cao nhất là hai giá trị đặc trưng của biên độ dao động ngẫu nhiên. Một số xảy ra gần trạng thái cân bằng, một số khác gần chu kỳ giới hạn, và có thể có sự chuyển đổi ít nhiều thường xuyên giữa các chế độ này. [...]
Các quần thể giả thuyết ứng xử theo các vectơ trong Hình. 10.11 A, được hiển thị trong hình. 10.11, -B với sự trợ giúp của biểu đồ thể hiện động thái của tỷ lệ số lượng vật ăn thịt và con mồi và trong hình. 10.11.5 dưới dạng đồ thị về sự biến động của số lượng vật ăn thịt và con mồi theo thời gian. Trong quần thể con mồi, khi nó chuyển từ trạng thái cân bằng mật độ thấp sang trạng thái cân bằng mật độ cao và quay trở lại, một sự "chớp nhoáng" về số lượng xảy ra. Và sự bùng phát này không phải là kết quả của một sự thay đổi rõ rệt trong môi trường. Ngược lại, sự thay đổi về số lượng này được tạo ra bởi chính tác động (với mức độ "ồn" thấp trong môi trường) và đặc biệt, nó phản ánh sự tồn tại của một số trạng thái cân bằng. Suy luận tương tự có thể được sử dụng để giải thích các trường hợp phức tạp hơn của biến động quần thể trong quần thể tự nhiên. […]
Đặc tính quan trọng nhất của hệ sinh thái là tính ổn định, sự cân bằng của trao đổi và các quá trình xảy ra trong đó. Khả năng của các quần thể hoặc hệ sinh thái để duy trì sự cân bằng động ổn định trong điều kiện môi trường thay đổi được gọi là cân bằng nội môi (homoios - giống nhau; stasis - trạng thái). Cân bằng nội môi dựa trên nguyên tắc phản hồi. Để duy trì sự cân bằng trong tự nhiên, không cần kiểm soát bên ngoài. Một ví dụ về cân bằng nội môi là hệ con "vật ăn thịt-con mồi", trong đó mật độ của quần thể vật ăn thịt và con mồi được quy định. [...]
Một hệ sinh thái tự nhiên (biogeocenosis) hoạt động ổn định với sự tương tác liên tục của các yếu tố của nó, sự tuần hoàn của các chất, sự chuyển giao năng lượng và thông tin hóa học, năng lượng, di truyền và năng lượng khác thông qua các kênh chuỗi. Theo nguyên tắc cân bằng, bất kỳ hệ thống tự nhiên nào có dòng năng lượng và thông tin đi qua nó đều có xu hướng phát triển một trạng thái ổn định. Đồng thời, sự ổn định của hệ sinh thái được cung cấp tự động do cơ chế phản hồi. Phản hồi bao gồm việc sử dụng dữ liệu nhận được từ các thành phần được quản lý của hệ sinh thái để thực hiện các điều chỉnh đối với các thành phần quản lý trong quy trình. Mối quan hệ "kẻ săn mồi" - "con mồi" được thảo luận ở trên trong bối cảnh này có thể được mô tả chi tiết hơn phần nào; vì vậy, trong hệ sinh thái thủy vực, cá săn mồi (chích chòe trong ao) ăn các loại cá săn mồi khác (cá diếc); nếu số lượng cá diếc sẽ tăng lên, đây là một ví dụ về phản hồi tích cực; pike, cho ăn cá diếc, làm giảm số lượng của nó - đây là một ví dụ về phản hồi tiêu cực; với sự gia tăng số lượng kẻ thù, số lượng nạn nhân giảm, và kẻ săn mồi, thiếu thức ăn, cũng làm giảm sự phát triển của dân số của nó; cuối cùng, trong ao đang được xem xét, sự cân bằng động được thiết lập với sự phong phú của cả cá chép và cá chép. Sự cân bằng được duy trì liên tục sẽ loại trừ sự biến mất của bất kỳ mắt xích nào trong chuỗi dinh dưỡng (Hình 64). [...]
Hãy chuyển sang phần khái quát quan trọng nhất, cụ thể là các tương tác tiêu cực trở nên ít được chú ý hơn theo thời gian nếu hệ sinh thái đủ ổn định và cấu trúc không gian của nó cho phép các quần thể điều chỉnh lẫn nhau. Trong hệ thống mô hình của kiểu động vật ăn thịt-con mồi, được mô tả bởi phương trình Lotka-Volterra, nếu các thuật ngữ bổ sung không được đưa vào phương trình đặc trưng cho ảnh hưởng của các yếu tố tự giới hạn của quần thể, thì sự biến động xảy ra liên tục và không chết (xem Levontin, 1969). Pimentel (1968; xem thêm Pimentel và Stone, 1968) đã chỉ ra bằng thực nghiệm rằng các thuật ngữ bổ sung như vậy có thể phản ánh sự thích nghi lẫn nhau hoặc phản hồi di truyền. Khi các nền văn hóa mới được tạo ra từ các cá thể trước đó đã cùng tồn tại trong một nền văn hóa trong hai năm, nơi mà số lượng của chúng phải chịu những biến động đáng kể, thì hóa ra chúng đã phát triển cân bằng nội môi sinh thái, trong đó mỗi quần thể bị “đàn áp” bởi quần thể kia ở mức độ mà nó hóa ra sự chung sống của họ ở một trạng thái cân bằng ổn định hơn.
Mô hình của Kolmogorov đưa ra một giả thiết quan trọng: vì người ta cho rằng điều này có nghĩa là có những cơ chế trong quần thể con mồi điều chỉnh sự phong phú của chúng ngay cả khi không có động vật ăn thịt.
Thật không may, việc xây dựng mô hình như vậy không cho phép chúng ta trả lời câu hỏi mà gần đây đã có rất nhiều tranh cãi và điều mà chúng ta đã đề cập ở đầu chương: làm thế nào một quần thể động vật ăn thịt có thể gây ra ảnh hưởng điều tiết lên con mồi dân số để toàn bộ hệ thống ổn định? Vì vậy, chúng ta sẽ quay trở lại mô hình (2.1), trong đó không có cơ chế tự điều chỉnh (ví dụ, điều chỉnh thông qua cạnh tranh nội bộ) trong quần thể con mồi (cũng như trong quần thể động vật ăn thịt); do đó, cơ chế duy nhất để điều chỉnh sự phong phú của các loài trong một quần xã là các mối quan hệ dinh dưỡng giữa vật ăn thịt và con mồi.
Ở đây (vì vậy, trái ngược với mô hình trước đó, tự nhiên các giải pháp (2.1) phụ thuộc vào kiểu chức năng sinh dưỡng cụ thể, do đó, được xác định bởi bản chất của sự săn mồi, tức là, chiến lược ăn thịt của kẻ săn mồi và chiến lược phòng thủ của con mồi. Chung cho tất cả các chức năng này (xem Hình I) là các đặc tính sau:
Hệ thống (2.1) có một điểm đứng yên bình thường có tọa độ được xác định từ các phương trình
![]()
với giới hạn tự nhiên.
Có thêm một điểm đứng yên (0, 0) ứng với trạng thái cân bằng. Dễ dàng chỉ ra rằng điểm này là yên ngựa, và các trục tọa độ là các đường phân cách.
Phương trình đặc trưng của một điểm có dạng
![]()
Rõ ràng, đối với mô hình Volterra cổ điển.
Do đó, giá trị của f có thể được coi là thước đo độ lệch của mô hình được xem xét so với mô hình Volterra.
![]()
điểm đứng yên là trọng tâm, trong hệ thống xuất hiện dao động; khi bất đẳng thức ngược lại được thỏa mãn, nó là một nút và không có dao động nào trong hệ. Sự ổn định của trạng thái cân bằng này được xác định bởi điều kiện
nghĩa là về cơ bản, nó phụ thuộc vào loại chức năng sinh dưỡng của động vật ăn thịt.
Điều kiện (5.5) có thể được hiểu như sau: đối với sự ổn định của trạng thái cân bằng tầm thường của hệ thống săn mồi - con mồi (và do đó đối với sự tồn tại của hệ thống này), chỉ cần tỷ lệ tương đối của con mồi được tiêu thụ là đủ. bởi động vật ăn thịt tăng lên cùng với sự gia tăng số lượng con mồi. Thật vậy, tỷ lệ con mồi (trong tổng số của chúng) được tiêu thụ bởi động vật ăn thịt được mô tả bằng một hàm có thể phân biệt được mà điều kiện tăng trưởng (tính dương của đạo hàm) trông như thế nào
![]()
Điều kiện cuối cùng, được thực hiện tại điểm, không gì khác ngoài điều kiện (5.5) của sự ổn định cân bằng. Với tính liên tục, nó cũng phải giữ ở một số vùng lân cận của điểm. Do đó, nếu số nạn nhân trong vùng lân cận này, thì
Bây giờ cho hàm dinh dưỡng V có dạng như trong Hình. 11a (đặc điểm của động vật không xương sống). Nó có thể được chỉ ra rằng với tất cả các giá trị hữu hạn (vì nó lồi lên trên)
tức là, bất bình đẳng (5.5) không được thỏa mãn đối với bất kỳ giá trị nào của số lượng nạn nhân cố định.
Điều này có nghĩa là trong một hệ thống có kiểu chức năng dinh dưỡng này không có trạng thái cân bằng ổn định không tầm thường. Một số kết quả có thể xảy ra: hoặc số lượng của cả con mồi và kẻ săn mồi tăng lên vô hạn, hoặc (khi quỹ đạo đi qua gần một trong các trục tọa độ), do những lý do ngẫu nhiên, số lượng con mồi hoặc số lượng kẻ săn mồi sẽ trở thành bằng không. Nếu con mồi chết, kẻ săn mồi sẽ chết sau một thời gian, nhưng nếu kẻ săn mồi chết trước, thì số lượng con mồi sẽ bắt đầu tăng lên theo cấp số nhân. Lựa chọn thứ ba - sự xuất hiện của một chu kỳ giới hạn ổn định - là không thể, điều này dễ dàng được chứng minh.
Thật vậy, biểu thức

trong góc phần tư dương luôn luôn dương, trừ khi nó có dạng như trong Hình. 11, a. Sau đó, theo tiêu chí của Dulac, không có quỹ đạo khép kín nào trong khu vực này và không thể tồn tại một chu kỳ giới hạn ổn định.
Vì vậy, chúng ta có thể kết luận: nếu chức năng dinh dưỡng có dạng như trong Hình. 11a, thì vật ăn thịt không thể là người điều tiết đảm bảo sự ổn định của quần thể con mồi và do đó sự ổn định của toàn bộ hệ thống nói chung. Hệ thống chỉ có thể ổn định nếu quần thể con mồi có các cơ chế điều tiết nội tại của riêng nó, chẳng hạn như cạnh tranh nội bộ hoặc động vật sinh sôi. Lựa chọn quy định này đã được xem xét trong §§ 3, 4.
Trước đây người ta đã lưu ý rằng kiểu chức năng dinh dưỡng này là đặc trưng của những kẻ săn mồi côn trùng, mà "nạn nhân" của chúng cũng thường là côn trùng. Mặt khác, các quan sát về động thái của nhiều quần xã tự nhiên thuộc kiểu “kẻ săn mồi”, bao gồm cả các loài côn trùng, cho thấy chúng có đặc điểm là dao động với biên độ rất lớn và một kiểu rất cụ thể.
Thông thường, sau khi số lượng tăng dần hoặc ít hơn (có thể xảy ra đơn điệu hoặc dưới dạng dao động với biên độ tăng dần), sự sụt giảm mạnh của nó xảy ra (Hình 14), và sau đó mô hình tự lặp lại. Rõ ràng, bản chất của động lực học của sự phong phú của các loài côn trùng có thể được giải thích bởi sự không ổn định của hệ thống này ở các giá trị phong phú thấp và trung bình và hoạt động của các cơ quan điều tiết mạnh mẽ của sự phong phú ở các giá trị lớn.

Cơm. Hình 14. Động thái quần thể của rầy chổng cánh Úc Cardiaspina albitextura ăn bạch đàn. (Từ bài báo: Clark L. R. Động thái dân số của Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, số 3, trang 362-380.)
Nếu hệ thống “động vật ăn thịt-con mồi” bao gồm các loài có khả năng hành vi khá phức tạp (ví dụ, động vật ăn thịt có khả năng học hỏi hoặc con mồi có thể tìm nơi trú ẩn), thì một hệ thống cân bằng ổn định không tầm thường có thể tồn tại. Việc chứng minh khẳng định này khá đơn giản.
Thật vậy, chức năng dinh dưỡng sau đó sẽ có dạng như trong Hình. 11, c. Điểm trên đồ thị là tiếp điểm của đường thẳng kẻ từ gốc tọa độ với đồ thị của hàm số có cực đại. Rõ ràng lúc này hàm số có cực đại. Cũng dễ dàng cho thấy rằng điều kiện (5.5) được thỏa mãn cho tất cả mọi người. Do đó, một trạng thái cân bằng không tầm thường trong đó số lượng nạn nhân ít hơn sẽ là tiệm cận ổn định
Tuy nhiên, chúng ta không thể nói bất cứ điều gì về vùng ổn định của trạng thái cân bằng này lớn như thế nào. Ví dụ, nếu có một chu kỳ giới hạn không ổn định, thì vùng này phải nằm bên trong chu trình. Hoặc một biến thể khác: trạng thái cân bằng không tầm thường (5.2) không ổn định, nhưng có một chu kỳ giới hạn ổn định; trong trường hợp này, người ta cũng có thể nói về sự ổn định của hệ thống săn mồi - con mồi. Kể từ biểu thức (5.7) khi chọn một chức năng dinh dưỡng như Hình. 11, dấu có thể thay đổi khi thay đổi tại, sau đó tiêu chí Dulac không hoạt động ở đây và câu hỏi về sự tồn tại của các chu kỳ giới hạn vẫn còn bỏ ngỏ.

Tương tác của các cá thể trong hệ thống "động vật ăn thịt-con mồi"
Sinh viên năm 5 51 Một nhóm
Khoa Sinh học
Nazarova A. A.
Cố vấn khoa học:
Podshivalov A. A.
Orenburg 2011
GIỚI THIỆU
GIỚI THIỆU
Trong các suy luận và quan sát hàng ngày của chúng ta, chúng ta, mà không tự mình biết và thậm chí không nhận ra nó, được hướng dẫn bởi các định luật và ý tưởng được khám phá từ nhiều thập kỷ trước. Xem xét vấn đề kẻ săn mồi - con mồi, chúng ta đoán rằng con mồi cũng ảnh hưởng gián tiếp đến kẻ săn mồi. Sư tử sẽ ăn gì nếu không có linh dương; người quản lý sẽ làm gì nếu không có công nhân; làm sao để phát triển kinh doanh nếu khách hàng không có kinh phí ...
Hệ thống "động vật ăn thịt-con mồi" là một hệ sinh thái phức tạp mà ở đó mối quan hệ lâu dài giữa động vật ăn thịt và con mồi được thực hiện, một ví dụ điển hình của hệ số tiến hóa. Mối quan hệ giữa động vật ăn thịt và con mồi của chúng phát triển theo chu kỳ, là một minh họa cho trạng thái cân bằng trung tính.
Việc nghiên cứu dạng quan hệ giữa các loài này, ngoài việc thu được những kết quả khoa học thú vị, còn cho phép chúng ta giải quyết nhiều vấn đề thực tế:
tối ưu hóa các biện pháp công nghệ sinh học cả trong mối quan hệ với các loài bắt mồi và mối quan hệ với động vật ăn thịt;
nâng cao chất lượng bảo vệ lãnh thổ;
quy định về áp lực săn bắn trong các trại săn bắn, v.v.
Những điều đã nói ở trên xác định mức độ liên quan của chủ đề đã chọn.
Mục đích của khóa học là nghiên cứu sự tương tác của các cá thể trong hệ thống "kẻ săn mồi - con mồi". Để đạt được mục tiêu, các nhiệm vụ sau đã được đặt ra:
sự săn mồi và vai trò của nó trong việc hình thành các mối quan hệ dinh dưỡng;
các mô hình chính của mối quan hệ "kẻ săn mồi - con mồi";
ảnh hưởng của lối sống xã hội đối với sự ổn định của hệ thống “kẻ săn mồi - con mồi”;
mô hình phòng thí nghiệm của hệ thống "kẻ săn mồi - con mồi".
Ảnh hưởng của động vật ăn thịt đối với số lượng con mồi và ngược lại là khá rõ ràng, nhưng khá khó để xác định cơ chế và thực chất của sự tương tác này. Những câu hỏi này tôi định giải quyết trong khóa học.
# ������� ############################################# ###### "# 5 # @ #? # 8 #; # 0 ### �� ##################### + ##### ###### �� \ ################ ################ �� # ��� ##### ######## Chương 4CHƯƠNG 4. MÔ HÌNH LAO ĐỘNG CỦA HỆ THỐNG TIÊN TIẾN - TRƯỚC
Các nhà khoa học của Đại học Duke, phối hợp với các đồng nghiệp từ Đại học Stanford, Viện Y khoa Howard Hughes và Viện Công nghệ California, làm việc dưới sự chỉ đạo của Tiến sĩ Lingchong You, đã phát triển một hệ thống sống của vi khuẩn biến đổi gen sẽ cho phép nghiên cứu chi tiết hơn. tương tác giữa vật ăn thịt và con mồi trong một quần thể.
Mô hình thử nghiệm mới là một ví dụ về hệ sinh thái nhân tạo, trong đó các nhà nghiên cứu lập trình vi khuẩn thực hiện các chức năng mới để tạo ra. Những vi khuẩn được lập trình lại như vậy có thể được sử dụng rộng rãi trong y học, làm sạch môi trường và phát triển máy tính sinh học. Là một phần của công trình này, các nhà khoa học đã viết lại "phần mềm" của E. coli (Escherichia coli) theo cách mà hai quần thể vi khuẩn khác nhau hình thành trong phòng thí nghiệm một hệ thống tương tác điển hình của động vật ăn thịt và con mồi, một đặc điểm của nó là vi khuẩn không ăn thịt lẫn nhau, nhưng kiểm soát số lượng dân số đối phương bằng cách thay đổi tần suất "tự sát".
Lĩnh vực nghiên cứu được gọi là sinh học tổng hợp xuất hiện vào khoảng năm 2000, và hầu hết các hệ thống được tạo ra kể từ đó đều dựa trên việc lập trình lại một loại vi khuẩn duy nhất. Mô hình do các tác giả phát triển độc đáo ở chỗ nó bao gồm hai quần thể vi khuẩn sống trong cùng một hệ sinh thái, sự tồn tại của chúng phụ thuộc vào nhau.
Chìa khóa cho sự hoạt động thành công của một hệ thống như vậy là khả năng của hai quần thể tương tác với nhau. Các tác giả đã tạo ra hai chủng vi khuẩn - "động vật ăn thịt" và "động vật ăn cỏ", tùy thuộc vào tình huống, giải phóng các hợp chất độc hại hoặc bảo vệ vào hệ sinh thái chung.
Nguyên tắc hoạt động của hệ thống dựa trên việc duy trì tỷ lệ giữa số lượng vật ăn thịt và con mồi trong môi trường quy định. Những thay đổi về số lượng tế bào ở một trong các quần thể sẽ kích hoạt các gen được lập trình lại, kích hoạt quá trình tổng hợp các hợp chất hóa học nhất định.
Do đó, một số ít nạn nhân trong môi trường gây ra sự kích hoạt của gen tự hủy trong tế bào động vật ăn thịt và cái chết của chúng. Tuy nhiên, khi số lượng nạn nhân tăng lên, hợp chất do họ thải ra môi trường đạt đến nồng độ tới hạn và kích hoạt gen động vật ăn thịt, đảm bảo tổng hợp "thuốc giải độc" cho gen tự sát. Điều này dẫn đến sự gia tăng dân số của những kẻ săn mồi, từ đó dẫn đến sự tích tụ của một hợp chất được tổng hợp bởi những kẻ săn mồi trong môi trường, đẩy nạn nhân đến chỗ tự sát.
Sử dụng kính hiển vi huỳnh quang, các nhà khoa học đã ghi lại những tương tác giữa động vật ăn thịt và con mồi.
Tế bào động vật ăn thịt, nhuộm xanh, làm tế bào con mồi tự sát, nhuộm đỏ. Sự kéo dài và vỡ của tế bào nạn nhân cho thấy nó đã chết.
Hệ thống này không phải là sự thể hiện chính xác các tương tác giữa động vật ăn thịt và con mồi trong tự nhiên, như vi khuẩn ăn thịt không ăn vi khuẩn bắt mồi và cả hai quần thể cạnh tranh để giành nguồn thức ăn như nhau. Tuy nhiên, các tác giả tin rằng hệ thống mà họ đã phát triển là một công cụ hữu ích cho nghiên cứu sinh học.
Hệ thống mới thể hiện mối quan hệ rõ ràng giữa di truyền và động lực học dân số, trong tương lai sẽ giúp nghiên cứu ảnh hưởng của các tương tác phân tử đối với sự thay đổi dân số, vốn là một chủ đề trung tâm của sinh thái học. Hệ thống cung cấp các khả năng gần như không giới hạn để sửa đổi các biến để nghiên cứu chi tiết các tương tác giữa môi trường, quy định gen và động lực của quần thể.
Do đó, bằng cách kiểm soát bộ máy di truyền của vi khuẩn, có thể mô phỏng các quá trình phát triển và tương tác của các sinh vật phức tạp hơn.
CHƯƠNG 3
CHƯƠNG 3
Các nhà sinh thái học từ Hoa Kỳ và Canada đã chỉ ra rằng lối sống theo nhóm của động vật ăn thịt và con mồi của chúng thay đổi hoàn toàn hành vi của hệ thống động vật ăn thịt và con mồi và làm cho nó linh hoạt hơn. Hiệu ứng này, được xác nhận bởi các quan sát về động thái của số lượng sư tử và linh dương đầu bò trong Công viên Serengeti, dựa trên thực tế đơn giản là với lối sống theo nhóm, tần suất chạm trán ngẫu nhiên giữa động vật ăn thịt và nạn nhân tiềm năng giảm.
Các nhà sinh thái học đã phát triển một số mô hình toán học mô tả hành vi của hệ thống động vật ăn thịt - con mồi. Đặc biệt, những mô hình này giải thích tốt những dao động định kỳ nhất quán đôi khi quan sát được về sự phong phú của các loài săn mồi và con mồi.
Các mô hình như vậy thường được đặc trưng bởi mức độ không ổn định cao. Nói cách khác, với một loạt các thông số đầu vào (chẳng hạn như tỷ lệ tử vong của động vật ăn thịt, hiệu quả chuyển đổi sinh khối của con mồi thành sinh khối của động vật ăn thịt, v.v.) trong các mô hình này, sớm hay muộn tất cả các động vật ăn thịt đều chết hoặc trước tiên là ăn hết con mồi, và sau đó vẫn chết vì đói.
Tất nhiên, trong các hệ sinh thái tự nhiên, mọi thứ phức tạp hơn so với trong một mô hình toán học. Rõ ràng, có nhiều yếu tố có thể làm tăng tính ổn định của hệ thống săn mồi - con mồi, và trên thực tế, hiếm khi xảy ra sự nhảy vọt về số lượng như ở linh miêu và thỏ rừng Canada.
Các nhà sinh thái học từ Canada và Hoa Kỳ đã công bố trong số mới nhất của tạp chí " thiên nhiên" một bài báo đã thu hút sự chú ý đến một yếu tố đơn giản và rõ ràng có thể thay đổi đáng kể hành vi của hệ thống săn mồi - con mồi. Đó là về cuộc sống nhóm.
Hầu hết các mô hình có sẵn đều dựa trên giả định về sự phân bố đồng đều của các loài săn mồi và con mồi của chúng trong một lãnh thổ nhất định. Đây là cơ sở để tính tần suất gặp gỡ của họ. Rõ ràng là mật độ con mồi càng cao, những kẻ săn mồi thường xuyên vấp phải chúng. Số lượng các cuộc tấn công, bao gồm cả những cuộc tấn công thành công, và cuối cùng, cường độ tấn công của những kẻ săn mồi phụ thuộc vào điều này. Ví dụ, với một con mồi dư thừa (nếu bạn không phải mất thời gian tìm kiếm), tốc độ ăn sẽ chỉ bị giới hạn trong khoảng thời gian cần thiết để kẻ săn mồi bắt, giết, ăn và tiêu hóa con mồi tiếp theo. Nếu con mồi hiếm khi bị bắt, yếu tố chính quyết định tốc độ ăn cỏ trở thành thời gian cần thiết để tìm kiếm con mồi.
Trong các mô hình sinh thái được sử dụng để mô tả hệ thống “kẻ săn mồi - con mồi”, bản chất của sự phụ thuộc của cường độ săn mồi (số lượng con mồi mà một kẻ săn mồi ăn trong một đơn vị thời gian) vào mật độ quần thể con mồi đóng một vai trò quan trọng. Loại thứ hai được ước tính là số lượng động vật trên một đơn vị diện tích.
Cần lưu ý rằng với lối sống theo nhóm gồm cả con mồi và động vật ăn thịt, giả thiết ban đầu về sự phân bố đồng đều trong không gian của các loài động vật là không thỏa mãn, và do đó tất cả các tính toán tiếp theo đều trở nên không chính xác. Ví dụ, với lối sống bầy đàn săn mồi, xác suất bắt gặp kẻ săn mồi thực sự sẽ không phụ thuộc vào số lượng cá thể động vật trên một km vuông, mà phụ thuộc vào số lượng đàn trên một đơn vị diện tích. Nếu con mồi được phân bổ đồng đều, những kẻ săn mồi sẽ tình cờ bắt gặp chúng thường xuyên hơn so với cách sống bầy đàn, vì những khoảng không rộng lớn được hình thành giữa bầy đàn, nơi không có con mồi. Một kết quả tương tự cũng thu được với cách sống theo nhóm của động vật ăn thịt. Niềm kiêu hãnh của những con sư tử lang thang trên thảo nguyên sẽ nhận thấy ít nạn nhân tiềm năng hơn là một con sư tử đơn độc đi theo con đường tương tự.
Trong ba năm (từ 2003 đến 2007), các nhà khoa học đã tiến hành quan sát kỹ lưỡng sư tử và nạn nhân của chúng (chủ yếu là linh dương đầu bò) trong lãnh thổ rộng lớn của Công viên Serengeti (Tanzania). Mật độ dân số được ghi nhận hàng tháng; cường độ ăn của sư tử thuộc các loài động vật móng guốc khác nhau cũng được đánh giá thường xuyên. Cả bản thân sư tử và bảy loài săn mồi chính của chúng đều có lối sống theo nhóm. Các tác giả đã đưa ra những sửa đổi cần thiết đối với các công thức sinh thái tiêu chuẩn để tính đến trường hợp này. Việc tham số hóa các mô hình được thực hiện trên cơ sở dữ liệu định lượng thực thu được trong quá trình quan sát. Bốn phiên bản của mô hình đã được xem xét: ở phiên bản đầu tiên, cách sống theo nhóm của động vật ăn thịt và con mồi bị bỏ qua; ở phiên bản thứ hai, nó chỉ được tính đến đối với động vật ăn thịt; ở phiên bản thứ ba, chỉ dành cho con mồi; và ở phiên bản thứ tư, cho cả hai.
|
|
Như người ta mong đợi, tùy chọn thứ tư phù hợp nhất với thực tế. Anh cũng tỏ ra là người kiên cường nhất. Điều này có nghĩa là với một loạt các thông số đầu vào trong mô hình này, có thể cùng tồn tại ổn định lâu dài giữa động vật ăn thịt và con mồi. Dữ liệu của các quan sát dài hạn cho thấy về mặt này, mô hình cũng phản ánh đầy đủ thực tế. Số lượng sư tử và con mồi của chúng ở Serengeti khá ổn định, không có gì giống với sự biến động phối hợp định kỳ (như trường hợp của linh miêu và thỏ rừng) được quan sát thấy.
Kết quả thu được cho thấy nếu sư tử và linh dương đầu bò sống đơn độc, số lượng con mồi tăng lên sẽ dẫn đến việc chúng bị kẻ thù săn mồi tăng tốc nhanh chóng. Do cách sống theo nhóm, điều này không xảy ra, hoạt động của những kẻ săn mồi tăng tương đối chậm, và mức độ săn mồi chung vẫn ở mức thấp. Theo các tác giả, được hỗ trợ bởi một số bằng chứng gián tiếp, số lượng nạn nhân ở Serengeti bị giới hạn không phải bởi sư tử, mà bởi nguồn thức ăn.
Nếu lợi ích của chủ nghĩa tập thể đối với các nạn nhân là khá rõ ràng, thì câu hỏi liên quan đến những con sư tử vẫn còn bỏ ngỏ. Nghiên cứu này đã chỉ ra rõ ràng rằng lối sống theo nhóm của những kẻ săn mồi có một nhược điểm nghiêm trọng - trên thực tế, vì nó, mỗi con sư tử cá thể kiếm được ít con mồi hơn. Rõ ràng, nhược điểm này cần được bù đắp bằng một số lợi thế rất đáng kể. Theo truyền thống, người ta tin rằng lối sống xã hội của sư tử gắn liền với việc săn bắt các loài động vật lớn, rất khó để đối phó ngay cả với một con sư tử. Tuy nhiên, gần đây, nhiều chuyên gia (bao gồm cả tác giả của bài báo đang thảo luận) bắt đầu nghi ngờ tính đúng đắn của lời giải thích này. Theo ý kiến của họ, hành động tập thể chỉ cần thiết đối với sư tử khi đi săn trâu, và sư tử thích đối phó với các loại con mồi khác một mình.
Hợp lý hơn là giả định rằng sự tự hào là cần thiết để điều chỉnh các vấn đề hoàn toàn bên trong, vốn có rất nhiều trong cuộc sống của một con sư tử. Ví dụ, trong số đó, tội giết người rất phổ biến - giết chết đàn con của người khác bởi con đực. Những con cái được nuôi trong một nhóm sẽ dễ dàng hơn để bảo vệ con cái của chúng khỏi những kẻ xâm lược. Ngoài ra, việc một con sư tử đơn độc bảo vệ khu vực săn mồi khỏi những con tự hào lân cận sẽ dễ dàng hơn rất nhiều đối với một niềm kiêu hãnh.
Nguồn: John M. Fryxell, Anna Mosser, Anthony R. E. Sinclair, Craig Packer. Sự hình thành nhóm ổn định động lực của kẻ săn mồi - con mồi // Thiên nhiên. 2007. V. 449. P. 1041–1043.
Mô phỏng hệ thống "Động vật ăn thịt-Nạn nhân"
Tóm tắt >> Mô hình kinh tế và toán học... hệ thống « Động vật ăn thịt-Nạn nhân" Thực hiện bởi Gizyatullin R.R gr.MP-30 Kiểm tra bởi Lisovets Yu.P MOSCOW 2007 Giới thiệu Sự tương tác... người mẫu tương tác kẻ săn mồi và nạn nhân trên bề mặt. Đơn giản hoá các giả định. Hãy thử so sánh nạn nhân và động vật ăn thịt một số...
Động vật ăn thịt-Nạn nhân
Tóm tắt >> Hệ sinh tháiCác ứng dụng của sinh thái toán học là hệ thống động vật ăn thịt-nạn nhân. Hành vi tuần hoàn của điều này hệ thống trong môi trường tĩnh là ... bằng cách đưa vào một mô hình phi tuyến tính bổ sung tương tác giữa động vật ăn thịt và một nạn nhân. Mô hình kết quả có trên ...
Tóm tắt nội dung sinh thái học
Tóm tắt >> Hệ sinh tháiyếu tố cho nạn nhân. Đó là lý do tại sao sự tương tác « động vật ăn thịt–nạn nhân" là định kỳ và là hệ thống Phương trình Lotka ... sự thay đổi nhỏ hơn nhiều so với trong hệ thống « động vật ăn thịt–nạn nhân". Tương tự tương tác cũng được quan sát thấy trong phép bắt chước Batsian. ...
Mô hình tương tác của hai loại
Các giả thuyết về Volterra. Tương tự với động học hóa học. Các mô hình tương tác của Volterra. Phân loại các loại tương tác Cạnh tranh. Động vật ăn thịt-con mồi. Các mô hình tương tác loài tổng quát . Mô hình Kolmogorov. Mô hình của MacArthur về sự tương tác giữa hai loài côn trùng. Tham số và chân dung pha của hệ thống Bazykin.
Nhà toán học người Ý Vito Volterra đúng ra được coi là người sáng lập ra lý thuyết toán học hiện đại về quần thể, người đã phát triển lý thuyết toán học về các cộng đồng sinh vật, bộ máy của nó là các phương trình vi phân và tích phân.(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris, Năm 1931). Trong những thập kỷ tiếp theo, động lực dân số phát triển chủ yếu phù hợp với những ý tưởng được trình bày trong cuốn sách này. Bản dịch tiếng Nga của cuốn sách của Volterra được xuất bản năm 1976 với tựa đề "Lý thuyết toán học về cuộc đấu tranh cho sự tồn tại" với lời bạt của Yu.M. Svirezhev, trong đó thảo luận về lịch sử phát triển của sinh thái toán học trong giai đoạn 1931-1976.
Cuốn sách của Volterra được viết theo cách những cuốn sách về toán học được viết. Đầu tiên nó hình thành một số giả định về các đối tượng toán học được cho là sẽ được nghiên cứu, và sau đó một nghiên cứu toán học về các thuộc tính của các đối tượng này được thực hiện.
Các hệ thống được Volterra nghiên cứu bao gồm hai hoặc nhiều loại. Trong một số trường hợp, dự trữ thực phẩm được sử dụng được xem xét. Các phương trình mô tả sự tương tác của các loài này dựa trên các biểu diễn sau.
Giả thuyết về Volterra
1. Thực phẩm có sẵn với số lượng không hạn chế, hoặc nguồn cung cấp theo thời gian được quản lý chặt chẽ.
2. Các cá thể của mỗi loài chết theo cách mà một tỷ lệ không đổi các cá thể hiện có bị chết trên một đơn vị thời gian.
3. Các loài săn mồi ăn con mồi, và trong một đơn vị thời gian số lượng con mồi ăn được luôn tỷ lệ thuận với xác suất gặp nhau của các cá thể của hai loài này, tức là sản phẩm của số lượng vật ăn thịt và số lượng con mồi.
4. Nếu có thức ăn với số lượng hạn chế và một số loài có khả năng tiêu thụ, thì tỷ lệ thức ăn của một loài tiêu thụ trong một đơn vị thời gian tỷ lệ với số lượng cá thể của loài này, lấy với một hệ số nhất định tùy thuộc vào các loài (mô hình cạnh tranh giữa các loài).
5. Nếu một loài ăn thức ăn có sẵn với số lượng không hạn chế, thì sự gia tăng số lượng của loài trên một đơn vị thời gian tỷ lệ thuận với số lượng của loài đó.
6. Nếu một loài ăn thức ăn có sẵn với số lượng hạn chế, thì khả năng sinh sản của chúng được điều chỉnh bởi tốc độ tiêu thụ thức ăn, tức là trên một đơn vị thời gian, sự gia tăng tỷ lệ thuận với lượng thức ăn ăn vào.
Tương tự với động học hóa học
Những giả thuyết này có sự tương đồng chặt chẽ với động học hóa học. Trong phương trình động lực học quần thể, cũng như trong phương trình động học hóa học, "nguyên tắc va chạm" được sử dụng, khi tốc độ phản ứng tỷ lệ với tích của nồng độ của các thành phần phản ứng.
Thật vậy, theo giả thuyết của Volterra, tốc độ quá trình sự tuyệt chủng của mỗi loài tỷ lệ thuận với sự phong phú của loài. Trong động học hóa học, điều này tương ứng với phản ứng phân hủy đơn phân tử của một số chất, và trong mô hình toán học, thành các số hạng tuyến tính âm ở vế phải của phương trình.
Theo khái niệm động học hóa học, tốc độ của một phản ứng lưỡng phân tử của sự tương tác của hai chất tỷ lệ thuận với xác suất va chạm của các chất này, tức là sản phẩm của nồng độ của chúng. Tương tự, theo giả thuyết của Volterra, tốc độ sinh sản của động vật ăn thịt (chết con mồi) tỷ lệ thuận với xác suất chạm trán giữa động vật ăn thịt và con mồi, tức là sản phẩm của những con số của họ. Trong cả hai trường hợp, các số hạng song tuyến xuất hiện trong hệ thống mô hình ở phía bên phải của các phương trình tương ứng.
Cuối cùng, các số hạng dương tuyến tính ở phía bên phải của phương trình Volterra, tương ứng với sự gia tăng dân số trong các điều kiện không bị hạn chế, tương ứng với các thuật ngữ tự xúc tác của phản ứng hóa học. Sự giống nhau của các phương trình trong mô hình hóa học và sinh thái làm cho nó có thể áp dụng các phương pháp nghiên cứu tương tự để mô hình hóa toán học về động học quần thể như đối với các hệ thống phản ứng hóa học.
Phân loại các loại tương tác
Theo giả thuyết của Volterra, sự tương tác của hai loài, số lượng x 1 và x 2 có thể được mô tả bằng các phương trình:
(9.1)
Đây là thông số một tôi - hằng số tốc độ tăng trưởng của các loài, c tôi- hằng số tự giới hạn dân số (cạnh tranh nội bộ cụ thể), b ij- hằng số tương tác giữa các loài, (tôi, j = 1,2). Dấu hiệu của các hệ số này xác định loại tương tác.
Trong tài liệu sinh học, các tương tác thường được phân loại theo các cơ chế liên quan. Sự đa dạng ở đây là rất lớn: các tương tác dinh dưỡng khác nhau, tương tác hóa học tồn tại giữa vi khuẩn và tảo phù du, tương tác của nấm với các sinh vật khác, sự kế tiếp của các sinh vật thực vật liên quan, đặc biệt, với sự cạnh tranh ánh sáng mặt trời và với sự tiến hóa của đất, v.v. Sự phân loại như vậy dường như không thể xác định được.
E . Odum, có tính đến các mô hình do V. Volterra đề xuất, đã đề xuất một cách phân loại không phải theo cơ chế, mà theo kết quả. Theo cách phân loại này, các mối quan hệ nên được đánh giá là tích cực, tiêu cực hoặc trung tính, tùy thuộc vào sự phong phú của một loài tăng, giảm hoặc không thay đổi khi có sự hiện diện của loài khác. Sau đó, các loại tương tác chính có thể được trình bày dưới dạng một bảng.
CÁC LOẠI TƯƠNG TÁC CỦA LOÀI
|
BIỂU TƯỢNG |
b 12 ,b 21 >0 |
||
|
GIAO LƯU |
b 12 ,>0, b 21 =0 |
||
|
PREDATOR-Con mồi |
b 12 ,>0, b 21 <0 |
||
|
AMENSALISM |
b 12 ,=0, b 21 <0 |
||
|
CUỘC ĐUA, CUỘC THI |
b 12 , b 21 <0 |
||
|
TRUNG LẬP |
b 12 , b 21 =0 |
Cột cuối cùng hiển thị các dấu hiệu của hệ số tương tác từ hệ thống (9.1)
Xem xét các loại tương tác chính
YÊU CẦU CẠNH TRANH:
Như chúng ta đã thấy trong Bài giảng 6, các phương trình cạnh tranh là:
 (9.2)
(9.2)
Giải pháp hệ thống văn phòng phẩm:
(1).
![]()
Gốc tọa độ, đối với bất kỳ tham số nào của hệ thống, là một nút không ổn định.
(2).
![]() (9.3)
(9.3)
C trạng thái đứng yên (9.3) là yên ngựa ở một 1 > b 12 /Với 2 và
nút thắt ổn định tại một 1 12 / s 2 . Điều kiện này có nghĩa là loài sẽ chết nếu tốc độ tăng trưởng của chúng thấp hơn một số giá trị quan trọng.
(3).
![]() (9.4)
(9.4)
C dung dịch tĩnh (9.4)¾ yên tại một 2 > b 21 /c 1 và một nút thắt ổn định ở một 2< b 21 /c 1
(4).
![]() (9.5)
(9.5)
Trạng thái tĩnh (9.5) đặc trưng cho sự cùng tồn tại của hai loài cạnh tranh và là một nút ổn định nếu mối quan hệ được thỏa mãn:
![]()
Điều này ngụ ý sự bất bình đẳng:
b 12
b 21
cho phép chúng ta hình thành điều kiện cho sự chung sống của các loài:
Tích của hệ số tương tác giữa các quần thể nhỏ hơn tích của các hệ số trong tương tác giữa quần thể.
Thật vậy, hãy để tốc độ sinh trưởng tự nhiên của hai loài được coi làmột 1 , một 2 giống nhau. Khi đó, điều kiện cần thiết để ổn định là
c 2 > b 12 ,c 1 > b 21 .
Những bất bình đẳng này cho thấy rằng sự gia tăng số lượng của một trong các đối thủ cạnh tranh kìm hãm sự phát triển của chính nó mạnh hơn sự tăng trưởng của một đối thủ cạnh tranh khác. Nếu sự phong phú của cả hai loài bị hạn chế, một phần hoặc toàn bộ, bởi các nguồn tài nguyên khác nhau, thì sự bất bình đẳng trên là có giá trị. Nếu cả hai loài đều có nhu cầu giống nhau, thì một trong số chúng sẽ khả thi hơn và sẽ thay thế đối thủ cạnh tranh của nó.
Hành vi của các quỹ đạo giai đoạn của hệ thống cung cấp một biểu thị trực quan về các kết quả có thể có của cạnh tranh. Ta cân bằng vế phải của các phương trình của hệ (9.2) bằng không:
x 1 (một 1 -c 1 x 1 – b 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (một 2 –b 21 x 1 – c 2 x 2) = 0 (dx 2 /dt = 0),
Trong trường hợp này, chúng ta thu được phương trình cho các đường đẳng chính của hệ thống
x 2 = - b 21 x 1 / c 2 +một 2 / c2, x 2 = 0
là phương trình của các đường đẳng của các tiếp tuyến thẳng đứng.
x 2 = - c 1 x 1 / b12 + một 1 / b 12 , x 1 = 0
là phương trình của các đường đẳng của các tiếp tuyến thẳng đứng. Các điểm giao nhau theo chiều kim của các đường đẳng của hệ tiếp tuyến dọc và ngang là nghiệm tĩnh của hệ phương trình (9.2.) Và tọa độ của chúng ![]() là số lượng cố định của các loài cạnh tranh.
là số lượng cố định của các loài cạnh tranh.
Vị trí có thể có của các đường đẳng chính trong hệ thống (9.2) được thể hiện trong Hình 9.1. Cơm. 9.1mộttương ứng với sự tồn tại của các loàix 1, hình 9.1 b- sự tồn tại của các loàix 2, hình 9.1 Trong- sự chung sống của các loài trong điều kiện (9.6). Hình 9.1Gtrình diễn hệ thống kích hoạt. Ở đây kết quả của cuộc thi phụ thuộc vào các điều kiện ban đầu. Trạng thái tĩnh (9.5), không phải là khác đối với cả hai loại, là không ổn định. Đây là cái yên ngựa mà ma trận đi qua, ngăn cách các khu vực sinh tồn của từng loài.

Cơm. 9.1.Vị trí của các đường đẳng chính trong chân dung pha của hệ thống Volterra cạnh tranh gồm hai loại (9.2) với các tỷ lệ thông số khác nhau. Các giải thích trong văn bản.
Để nghiên cứu sự cạnh tranh của các loài, các thí nghiệm đã được thực hiện trên nhiều loại sinh vật. Thông thường, hai loài có quan hệ họ hàng gần được lựa chọn và trồng cùng nhau và riêng biệt trong các điều kiện được kiểm soát nghiêm ngặt. Vào những khoảng thời gian nhất định, một cuộc tổng điều tra dân số hoàn chỉnh hoặc có chọn lọc được thực hiện. Ghi lại dữ liệu từ một số thí nghiệm lặp lại và phân tích. Các nghiên cứu được thực hiện trên động vật nguyên sinh (đặc biệt là ciliates), nhiều loài bọ cánh cứng thuộc chi Tribolium, Drosophila và giáp xác nước ngọt (daphnia). Nhiều thí nghiệm đã được thực hiện trên các quần thể vi sinh vật (xem bài giảng 11). Các thí nghiệm cũng được thực hiện trong tự nhiên, bao gồm cả trên người phẳng (Reynolds), hai loài kiến (Pontin) và những loài khác. 9.2. các đường cong tăng trưởng của tảo cát sử dụng cùng một nguồn tài nguyên (chiếm cùng một ngách sinh thái) được hiển thị. Khi trồng độc canh Asterionella formosa đạt đến mức mật độ không đổi và duy trì nồng độ của tài nguyên (silicat) ở mức thấp liên tục. B. Khi trồng độc canh Synedrauina hoạt động theo cách tương tự và giữ nồng độ silicat ở mức thấp hơn. B. Cùng canh tác (nhân đôi) Synedrauina vượt qua bệnh Asterionella formosa. Rõ ràng là Synedra

Cơm. 9.2.Cạnh tranh trong tảo cát. một - khi trồng trong độc canh Asterionella formosa đạt đến mức mật độ không đổi và duy trì nồng độ của tài nguyên (silicat) ở mức thấp liên tục. b - khi trồng trong độc canh Synedrauina hoạt động theo cách tương tự và giữ nồng độ silicat ở mức thấp hơn. Trong - trong cùng trồng trọt (trùng lặp) Synedruina vượt qua bệnh Asterionella formosa. Rõ ràng là Synedra thắng cuộc cạnh tranh do khả năng sử dụng đầy đủ hơn chất nền (xem thêm Bài giảng 11).
Các thí nghiệm của G. Gause về nghiên cứu cạnh tranh được biết đến rộng rãi, chứng minh sự sống còn của một trong những loài cạnh tranh và cho phép ông xây dựng "luật loại trừ cạnh tranh". Luật quy định rằng chỉ có một loài có thể tồn tại trong một vùng sinh thái. Trên hình. 9.3. Kết quả của các thí nghiệm của Gause đối với hai loài Parametium chiếm cùng một ngách sinh thái (Hình 9.3 a, b) và các loài chiếm các hốc sinh thái khác nhau (Hình 9.3. c) được trình bày.

Cơm. 9.3. một- Đường cong tăng trưởng quần thể của hai loài Parametium trong các nền văn hóa đơn loài. Vòng tròn đen - P Aurelia, vòng tròn trắng - P. Caudatum
b- Đường cong tăng trưởng P aurelia và P. Caudatum trong một nền văn hóa hỗn hợp.
Bởi Gause, 1934
Mô hình cạnh tranh (9.2) có những thiếu sót, cụ thể là nó cho rằng sự chung sống của hai loài chỉ có thể thực hiện được nếu mức độ phong phú của chúng bị hạn chế bởi các yếu tố khác nhau, nhưng mô hình không chỉ ra sự khác biệt phải lớn đến mức nào để đảm bảo sự chung sống lâu dài. . Đồng thời, biết rằng cùng tồn tại lâu dài trong môi trường thay đổi đòi hỏi sự khác biệt đạt đến một giá trị nhất định. Việc đưa các yếu tố ngẫu nhiên vào mô hình (ví dụ, giới thiệu một hàm sử dụng tài nguyên) cho phép chúng ta nghiên cứu một cách định lượng các vấn đề này.
Hệ thống săn mồi + con mồi
 (9.7)
(9.7)
Ở đây, trái ngược với (9.2), các dấu hiệu b 12 và b 21 - khác nhau. Như trong trường hợp cạnh tranh, nguồn gốc
![]() (9.8)
(9.8)
là một điểm số ít của loại nút không ổn định. Ba trạng thái tĩnh có thể có khác:
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
Vì vậy, chỉ có con mồi (9.10), chỉ có động vật ăn thịt (9.9) (nếu nó có nguồn thức ăn khác) và sự chung sống của cả hai loài (9.11) là có thể. Phương án cuối cùng đã được chúng tôi xem xét trong bài giảng 5. Các kiểu chân dung pha có thể có đối với hệ thống săn mồi-con mồi được trình bày trong Hình. 9.4.
Đường đẳng của các tiếp tuyến nằm ngang là các đường thẳng
x 2 = – b 21 X 1 /c 2 + một 1 / c2, X 2 = 0,
và các đường đẳng của các tiếp tuyến thẳng đứng- dài
x 2 = - c 1 X 1 /b 12 + một 2 /b 12 , X 1 = 0.
Các điểm đứng yên tại giao điểm của các đường đẳng của các tiếp tuyến dọc và ngang.

Từ hình 9.4 sau đây được nhìn thấy. hệ thống săn mồi-con mồi (9.7) có thể có một vị trí cân bằng ổn định, trong đó o rum, quần thể nạn nhân đã hoàn toàn tuyệt chủng ( ) và chỉ còn lại những kẻ săn mồi (dấu chấm 2 trong hình. 9.4 một). Rõ ràng, tình huống như vậy chỉ có thể thành hiện thực nếu, ngoài loại nạn nhân đang được xem xét, X 1 kẻ săn mồi X 2 - có nguồn điện bổ sung. Thực tế này được phản ánh trong mô hình bằng số hạng dương ở vế phải của phương trình đối với x 2. Điểm số ít(1) và (3) (Hình 9.4 một) không ổn định. Khả năng thứ hai – một trạng thái tĩnh ổn định trong đó quần thể động vật ăn thịt đã chết hoàn toàn và chỉ còn lại nạn nhân – điểm ổn định(3) (Hình 9.4 6 ). Đây là một điểm đặc biệt (1) – cũng là một nút không ổn định.
Cuối cùng, khả năng thứ ba – sự chung sống ổn định của quần thể động vật ăn thịt và con mồi (Hình. 9.4 Trong), có độ dồi dào tĩnh tại được biểu thị bằng công thức (9.11).
Như trong trường hợp của một tập hợp đơn lẻ (xem Bài giảng 3), đối với mô hình (9.7) nó có thể phát triển một mô hình ngẫu nhiên, nhưng nó không thể được giải quyết một cách rõ ràng. Do đó, chúng ta tự giới hạn mình trong những cân nhắc chung chung. Ví dụ, giả sử rằng điểm cân bằng nằm ở một khoảng cách nào đó từ mỗi trục. Sau đó, đối với quỹ đạo pha mà các giá trịx 1 , x 2 vẫn đủ lớn, một mô hình xác định sẽ khá thỏa đáng. Nhưng nếu tại một thời điểm nào đó của quỹ đạo pha một biến số nào đó không lớn lắm, thì các dao động ngẫu nhiên có thể trở nên đáng kể. Chúng dẫn đến thực tế là điểm đại diện sẽ di chuyển đến một trong các trục, đồng nghĩa với sự tuyệt chủng của các loài tương ứng.
Do đó, mô hình ngẫu nhiên hóa ra không ổn định, vì sự “trôi dạt” ngẫu nhiên sớm hay muộn dẫn đến sự tuyệt chủng của một trong các loài. Trong loại mô hình này, động vật ăn thịt cuối cùng chết đi, do ngẫu nhiên hoặc do quần thể con mồi của nó bị loại bỏ trước. Mô hình ngẫu nhiên của hệ thống săn mồi-con mồi giải thích tốt các thí nghiệm của Gause (Gause, 1934), trong đó các chi nhánh Paramettum candatum phục vụ như một con mồi cho một con ciliate khác Didinium nasatum – động vật ăn thịt. Dự kiến theo phương trình xác định (9.7) số lượng cân bằng trong các thí nghiệm này chỉ xấp xỉ năm cá thể của mỗi loài, vì vậy không có gì đáng ngạc nhiên trong thực tế là trong mỗi thí nghiệm lặp lại, động vật ăn thịt hoặc con mồi (và sau đó là động vật ăn thịt) chết khá nhanh. trong bộ lễ phục. 9,5.

Cơm. 9.5. Sự phát triển Parametium caudatum và các chi nhánh săn mồi Dadinium mũi họng. Từ : Vì G.F. Cuộc đấu tranh cho sự tồn tại. Baltimore, 1934
Vì vậy, việc phân tích các mô hình Volterra về sự tương tác giữa các loài cho thấy rằng, mặc dù có rất nhiều kiểu hành vi của các hệ thống như vậy, nhưng không thể có sự biến động quần thể nào chưa được đánh dấu trong mô hình của các loài cạnh tranh. Tuy nhiên, những dao động như vậy được quan sát trong tự nhiên và trong thực nghiệm. Nhu cầu giải thích lý thuyết của họ là một trong những lý do để xây dựng mô tả mô hình ở dạng tổng quát hơn.
Mô hình tổng quát về sự tương tác của hai loại
Một số lượng lớn các mô hình đã được đề xuất mô tả sự tương tác của các loài, vế phải của các phương trình là hàm của kích thước của các quần thể tương tác. Vấn đề xây dựng các tiêu chí chung để xác định loại chức năng nào có thể mô tả hành vi của quy mô dân số tạm thời, bao gồm cả những biến động ổn định, đã được xem xét. Nổi tiếng nhất trong số các mô hình này là của Kolmogorov (1935, sửa đổi 1972) và Rosenzweig (1963).
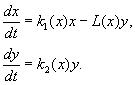 (9.12)
(9.12)
Mô hình dựa trên các giả định sau:
1) Động vật ăn thịt không tương tác với nhau, tức là tỷ lệ sinh sản của động vật ăn thịt k 2 và số nạn nhân L, bị tiêu diệt trên một đơn vị thời gian bởi một kẻ săn mồi, không phụ thuộc vào y.
2) Sự gia tăng số lượng con mồi khi có vật ăn thịt bằng sự gia tăng khi không có vật ăn thịt trừ đi số lượng con mồi bị tiêu diệt bởi vật ăn thịt. Chức năng k 1 (x), k 2 (x), L(x), liên tục và được xác định trên bán trục dương x, y³ 0.
3) dk 1 / dx< 0. Điều này có nghĩa là hệ số nhân lên của con mồi khi không có động vật ăn thịt sẽ giảm đi một cách đơn điệu khi số lượng con mồi tăng lên, điều này phản ánh sự hạn chế của thức ăn và các nguồn tài nguyên khác.
4) dk 2 / dx> 0, k 2 (0) < 0 < k 2 (¥ ). Với sự gia tăng số lượng con mồi, hệ số nhân của động vật ăn thịt giảm đơn điệu khi số lượng con mồi tăng lên, chuyển từ giá trị âm (khi không có gì để ăn) sang giá trị dương.
5) Số lượng nạn nhân bị tiêu diệt bởi một kẻ săn mồi trên một đơn vị thời gian L(x)> 0 tại N> 0; L(0)=0.
Các kiểu chân dung pha có thể có của hệ thống (9.12) được trình bày trong hình. 9,6:

Cơm. 9,6.Chân dung pha của hệ thống Kolmogorov (9.12), mô tả sự tương tác của hai loại đối với các tỷ lệ tham số khác nhau. Các giải thích trong văn bản.
Các nghiệm tĩnh (có hai hoặc ba trong số chúng) có tọa độ sau:
(1). ` x = 0;` y = 0.
Gốc tọa độ cho bất kỳ giá trị nào của các tham số là một cái yên (Hình 9.6 a-d).
(2). ` x = A,` y = 0.(9.13)
Mộtđược xác định từ phương trình:
k 1 (Một)=0.
Đứng im giải pháp (9.13) là yên nếu B< Một (Hình 9.6 một, b, G), B xác định từ phương trình
k 2 (B)=0
Điểm (9.13) được đặt trong góc phần tư dương nếu B> A . Đây là một nút thắt ổn định .
Trường hợp cuối cùng, tương ứng với cái chết của kẻ săn mồi và sự sống sót của con mồi, được thể hiện trong Hình. 9.6 Trong.
(3). ` x = B,` y = C.(9.14)
Giá trị của C được xác định theo phương trình:
Điểm (9.14) - tiêu điểm (Hình 9.6 một) hoặc nút (Hình 9.6 G), độ ổn định của nó phụ thuộc vào dấu hiệu của số lượngS
S 2 = – k 1 (B) -k 1 (B)B + L(B)C.
Nếu một S>0, điểm ổn định nếuS<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 b)
Trong tài liệu nước ngoài, một mô hình tương tự do Rosenzweig và MacArthur (1963) đề xuất thường được xem xét nhiều hơn:
 (9.15)
(9.15)
ở đâu f(x) - tỷ lệ thay đổi số lượng nạn nhân x trong trường hợp không có động vật ăn thịt, F ( x, y) là cường độ của sự săn mồi, k- hệ số đặc trưng cho hiệu quả chuyển đổi sinh khối con mồi thành sinh khối vật ăn thịt, e- Tỷ lệ tử vong của động vật ăn thịt.
Mô hình (9.15) rút gọn thành một trường hợp cụ thể của mô hình Kolmogorov (9.12) theo các giả định sau:
1) số lượng động vật ăn thịt chỉ bị giới hạn bởi số lượng con mồi,
2) Tốc độ một cá thể của động vật ăn thịt nhất định chỉ phụ thuộc vào mật độ quần thể con mồi và không phụ thuộc vào mật độ quần thể động vật ăn thịt.
Khi đó phương trình (9.15) có dạng.

Khi mô tả sự tương tác của các loài thực, các phần bên phải của phương trình được cụ thể hóa phù hợp với các ý tưởng về thực tại sinh học. Hãy xem xét một trong những mô hình phổ biến nhất của loại này.
Mô hình tương tác giữa hai loài côn trùng (MacArthur, 1971)
Mô hình mà chúng ta sẽ thảo luận dưới đây, được sử dụng để giải quyết vấn đề thực tế về kiểm soát dịch hại bằng cách khử trùng con đực của một trong các loài. Dựa trên các đặc điểm sinh học của sự tương tác giữa các loài, mô hình sau đây đã được viết
 (9.16)
(9.16)
Nơi đây x, y- sinh khối của hai loài côn trùng. Tương tác dinh dưỡng của các loài được mô tả trong mô hình này rất phức tạp. Điều này xác định dạng của đa thức ở vế phải của phương trình.
Xét vế phải của phương trình đầu tiên. Loài côn trùng Xăn ấu trùng của loài tại(thành viên + k 3 y), nhưng người lớn của loài tạiăn ấu trùng của loài X tùy thuộc vào số lượng loài cao X hoặc tại hoặc cả hai loại (thành viên –K 4 xy, - y 2). Nhỏ X tỷ lệ chết của các loài X cao hơn mức tăng tự nhiên của nó (1 –K 1 + k 2 x – x 2 < 0 nhỏ X). Trong phương trình thứ hai, thuật ngữ k 5 phản ánh sự phát triển tự nhiên của các loài y; –K 6 y- sự tự kiềm chế của loại này,–K 7 x- ăn ấu trùng của loài tại côn trùng của các loài x, k 8 xy – tăng trưởng sinh khối loài tại bị ăn bởi côn trùng trưởng thành của loài tạiấu trùng của các loài X.
Trên hình. 9,7 chu trình giới hạn được trình bày, là quỹ đạo của một dung dịch tuần hoàn ổn định của hệ thống (9.16).

Tất nhiên, không thể có lời giải cho câu hỏi làm thế nào để đảm bảo sự chung sống của một quần thể với môi trường sinh học của nó nếu không tính đến các đặc điểm cụ thể của một hệ thống sinh học cụ thể và phân tích tất cả các mối tương quan của nó. Đồng thời, việc nghiên cứu các mô hình toán học chính thức giúp chúng ta có thể trả lời một số câu hỏi tổng quát. Có thể lập luận rằng đối với các mô hình thuộc loại (9.12), sự tương thích hay không tương thích của các quần thể không phụ thuộc vào kích thước ban đầu của chúng, mà chỉ được xác định bởi bản chất của sự tương tác giữa các loài. Mô hình giúp trả lời câu hỏi: làm thế nào để tác động đến hiện tượng vi sinh vật, quản lý nó để tiêu diệt các loài gây hại càng nhanh càng tốt.
Quản lý có thể được giảm xuống một sự thay đổi ngắn hạn, co thắt về quy mô dân số X và y. Phương pháp này tương ứng với các phương pháp kiểm soát như tiêu diệt đơn lẻ một hoặc cả hai quần thể bằng biện pháp hóa học. Từ tuyên bố được xây dựng ở trên, có thể thấy rằng đối với các quần thể tương thích, phương pháp kiểm soát này sẽ không hiệu quả, vì theo thời gian hệ thống sẽ lại chuyển sang chế độ cố định.
Một cách khác là thay đổi kiểu hàm tương tác giữa các kiểu, ví dụ khi thay đổi giá trị của các tham số hệ thống. Chính phương pháp tham số này tương ứng với các phương pháp đấu tranh sinh học. Như vậy, khi đưa những con đực đã triệt sản vào thì hệ số gia tăng dân số tự nhiên giảm xuống. Nếu đồng thời chúng ta nhận được một loại chân dung pha khác, một loại chỉ có trạng thái tĩnh ổn định với số sâu bệnh bằng không, thì việc kiểm soát sẽ dẫn đến kết quả mong muốn – tiêu diệt quần thể dịch hại. Điều thú vị cần lưu ý là đôi khi chúng ta nên tác động không phải đối với dịch hại mà là đối với đối tác của chúng. Phương pháp nào hiệu quả hơn, trong trường hợp chung, không thể nói. Nó phụ thuộc vào các biện pháp kiểm soát có sẵn và vào dạng rõ ràng của các chức năng mô tả sự tương tác của các quần thể.
Người mẫu A.D.Bazykin
Phân tích lý thuyết về các mô hình tương tác giữa các loài được thực hiện đầy đủ nhất trong cuốn sách của A.D. Bazykin “Lý sinh học của các quần thể tương tác” (M., Nauka, 1985).
Hãy xem xét một trong những mô hình kẻ săn mồi - con mồi được nghiên cứu trong cuốn sách này.
 (9.17)
(9.17)
Hệ thống (9.17) là sự khái quát hóa mô hình động vật ăn thịt - con mồi Volterra đơn giản nhất (5.17) có tính đến hiệu ứng bão hòa của động vật ăn thịt. Mô hình (5.17) giả định rằng cường độ săn mồi tăng tuyến tính khi mật độ con mồi tăng, điều này không tương ứng với thực tế ở mật độ con mồi cao. Các chức năng khác nhau có thể được chọn để mô tả sự phụ thuộc của khẩu phần ăn của động vật ăn thịt vào mật độ con mồi. Điều quan trọng nhất là chức năng được chọn với sự gia tăng x có xu hướng tiệm cận với một giá trị không đổi. Mô hình (9.6) đã sử dụng sự phụ thuộc vào logistic. Trong mô hình Bazykin, hyperbola được chọn làm một hàm như vậy x/(1+ px). Nhớ lại rằng công thức của Monod, mô tả sự phụ thuộc của tốc độ phát triển của vi sinh vật vào nồng độ của chất nền, có dạng này. Ở đây, con mồi hoạt động như một chất nền, và động vật ăn thịt đóng vai trò như vi sinh vật. .
Hệ thống (9.17) phụ thuộc vào bảy tham số. Có thể giảm số lượng tham số bằng cách thay đổi các biến:
x® (A / D)x; y ® (A / D)/ y;
t® (1 / A)t; g (9,18)
và phụ thuộc vào bốn tham số.
Để có một nghiên cứu định tính hoàn chỉnh, cần phải chia không gian tham số bốn chiều thành các vùng có các kiểu hành vi động khác nhau, tức là xây dựng chân dung tham số hoặc cấu trúc của hệ thống.
Sau đó, cần phải xây dựng chân dung pha cho từng vùng của chân dung tham số và mô tả sự phân đôi xảy ra với chân dung theo pha tại ranh giới của các vùng khác nhau của chân dung tham số.
Việc xây dựng một chân dung tham số hoàn chỉnh được thực hiện dưới dạng một tập hợp các “lát cắt” (hình chiếu) của một chân dung tham số có kích thước nhỏ với các giá trị cố định của một số tham số.
Chân dung tham số của hệ thống (9.18) cho cố định g và nhỏ eđược thể hiện trong Hình 9.8. Bức chân dung chứa 10 khu vực với các loại hành vi quỹ đạo pha khác nhau.

Cơm. 9,8.Chân dung tham số của hệ thống (9.18) cho cố địnhg
và nhỏ e
Hành vi của hệ thống với các tỷ lệ thông số khác nhau có thể khác nhau đáng kể (Hình 9.9). Những điều sau có thể có trong hệ thống:
1) một trạng thái cân bằng ổn định (vùng 1 và 5);
2) một chu kỳ giới hạn ổn định (vùng 3 và 8);
3) hai điểm cân bằng ổn định (vùng 2)
4) chu kỳ giới hạn ổn định và trạng thái cân bằng không ổn định bên trong nó (vùng 6, 7, 9, 10)
5) chu kỳ giới hạn ổn định và cân bằng ổn định bên ngoài nó (vùng 4).
Trong các vùng tham số 7, 9, 10, vùng cân bằng lực hút bị giới hạn bởi một chu kỳ giới hạn không ổn định nằm bên trong chu trình ổn định. Thú vị nhất là chân dung pha tương ứng với vùng 6 trong chân dung tham số. Nó được thể hiện chi tiết trong Hình. 9,10.

Vùng hấp dẫn của trạng thái cân bằng B 2 (được tô bóng) là một "con ốc" xoắn khỏi tiêu điểm không ổn định B 1. Nếu biết rằng tại thời điểm ban đầu hệ đang ở lân cận В 1 thì có thể phán đoán quỹ đạo tương ứng sẽ tiến tới cân bằng В 2 hay đến một chu kỳ giới hạn ổn định xung quanh ba điểm cân bằng С ( yên ngựa), В 1 và В 2 chỉ dựa trên các cân nhắc xác suất.

Hình.9.10.Chân dung pha của hệ 9.18 cho vùng tham số 6. Vùng hấp dẫn B 2 được tô bóng
Trên một bức chân dung tham số(9,7) có 22 các ranh giới phân đôi khác nhau hình thành 7 các loại phân nhánh khác nhau. Nghiên cứu của họ giúp xác định các loại hành vi có thể có của hệ thống khi các tham số của nó thay đổi. Ví dụ, khi di chuyển từ khu vực 1 đến khu vực 3 có sự sinh ra của một chu kỳ giới hạn nhỏ, hoặc sự sinh ra của các dao động tự động quanh một điểm cân bằng duy nhất. TẠI. Một sự sinh ra mềm tương tự của các dao động tự, nhưng xung quanh một trong những điểm cân bằng, cụ thể là B 1 , xảy ra khi đi qua biên giới của các vùng 2 và 4. Khi di chuyển từ khu vực 4 đến khu vực 5 chu kỳ giới hạn ổn định xung quanh một điểmB 1 "Bùng nổ" trên vòng lặp phân tách và điểm thu hút duy nhất là điểm cân bằng B 2 vân vân.
Tất nhiên, mối quan tâm đặc biệt đối với thực tiễn là việc phát triển các tiêu chí cho sự gần gũi của một hệ thống với các ranh giới phân đôi. Thật vậy, các nhà sinh vật học nhận thức rõ về tính chất "đệm" hay "tính linh hoạt" của các hệ thống sinh thái tự nhiên. Các thuật ngữ này thường biểu thị khả năng của hệ thống để hấp thụ các tác động bên ngoài, như nó vốn có. Miễn là cường độ của hành động bên ngoài không vượt quá một giá trị tới hạn nhất định, hành vi của hệ thống không trải qua những thay đổi về chất. Trên mặt phẳng pha, điều này tương ứng với việc hệ trở lại trạng thái cân bằng ổn định hoặc về một chu kỳ giới hạn ổn định, các thông số của chúng không khác nhiều so với ban đầu. Khi cường độ của tác động vượt quá mức cho phép, hệ thống sẽ "phá vỡ", chuyển sang một chế độ hành vi động khác về chất, chẳng hạn, nó chỉ đơn giản là chết đi. Hiện tượng này tương ứng với sự chuyển đổi phân đôi.
Mỗi loại chuyển đổi phân nhánh đều có những đặc điểm nổi bật riêng để có thể đánh giá mức độ nguy hiểm của quá trình chuyển đổi như vậy đối với hệ sinh thái. Dưới đây là một số tiêu chí chung chứng minh sự gần kề của ranh giới nguy hiểm. Như trong trường hợp của một loài, nếu sự giảm số lượng của một trong số các loài khiến hệ thống "mắc kẹt" ở gần điểm yên ngựa không ổn định, được thể hiện ở việc số lượng phục hồi rất chậm về giá trị ban đầu, thì hệ thống ở gần ranh giới tới hạn. Sự thay đổi dạng dao động của số lượng kẻ săn mồi và con mồi cũng là một dấu hiệu cho thấy sự nguy hiểm. Nếu các dao động trở nên giãn từ gần điều hòa và biên độ dao động tăng lên, điều này có thể dẫn đến sự mất ổn định của hệ thống và sự tuyệt chủng của một trong các loài.
Việc đào sâu hơn nữa lý thuyết toán học về sự tương tác của các loài đi cùng với con đường chi tiết hóa cấu trúc của các quần thể và tính đến các yếu tố thời gian và không gian.
Văn chương.
Kolmogorov A.N. Nghiên cứu định tính các mô hình toán học về động lực dân số. // Các vấn đề về điều khiển học. M., 1972, số 5.
MacArtur R. Phân tích đồ thị các hệ thống sinh thái // Báo cáo bộ phận sinh học Đại học Perinceton. 1971
AD Bazykin "Lý sinh của các quần thể tương tác". M., Nauka, 1985.
W. Volterra: "Lý thuyết toán học về cuộc đấu tranh cho sự tồn tại." M.. Khoa học, 1976
Gạc G.F. Cuộc đấu tranh cho sự tồn tại. Baltimore, năm 1934.
Ở đây, trái ngược với (3.2.1), các dấu hiệu (-012) và (+ a2i) là khác nhau. Như trong trường hợp cạnh tranh (hệ phương trình (2.2.1)), điểm gốc (1) của hệ này là một điểm kỳ dị của kiểu “nút không ổn định”. Ba trạng thái tĩnh có thể có khác:

Ý nghĩa sinh học đòi hỏi những giá trị tích cực X y x 2. Đối với biểu thức (3.3.4), điều này có nghĩa là
Nếu hệ số cạnh tranh nội bộ của các loài săn mồi một,22 = 0, điều kiện (3.3.5) dẫn đến điều kiện ai2
Các dạng chân dung pha có thể có đối với hệ phương trình (3.3.1) được trình bày trong hình. 3.2 a-c. Đường đẳng của các tiếp tuyến nằm ngang là các đường thẳng
và các đường đẳng của các tiếp tuyến thẳng đứng là đường thẳng
Từ hình 3.2 cho thấy những điều sau đây. Hệ thống săn mồi - con mồi (3.3.1) có thể có trạng thái cân bằng ổn định trong đó quần thể con mồi hoàn toàn tuyệt chủng (x = 0) và chỉ còn lại những kẻ săn mồi (điểm 2 trong Hình 3.26). Rõ ràng, tình huống như vậy chỉ có thể thành hiện thực nếu, ngoài loại nạn nhân đang được xem xét, Xđộng vật ăn thịt X2 có bộ nguồn bổ sung. Thực tế này được phản ánh trong mô hình bằng số hạng dương ở vế phải của phương trình đối với xs. Các điểm số ít (1) và (3) (Hình 3.26) không ổn định. Khả năng thứ hai là trạng thái tĩnh tại ổn định, trong đó quần thể động vật ăn thịt chết hoàn toàn và chỉ còn lại nạn nhân - một điểm ổn định (3) (Hình 3.2a). Ở đây điểm kỳ dị (1) cũng là một nút không ổn định.
Cuối cùng, khả năng thứ ba là sự chung sống ổn định của các quần thể động vật ăn thịt và con mồi (Hình 3.2 c), chúng có số lượng đứng yên được biểu thị bằng công thức (3.3.4). Chúng ta hãy xem xét trường hợp này chi tiết hơn.
Giả sử rằng các hệ số của cạnh tranh nội bộ cụ thể bằng 0 (ai= 0, i = 1, 2). Chúng ta cũng hãy giả định rằng những kẻ săn mồi chỉ ăn con mồi của loài X và khi không có chúng, chúng chết đi với tỷ lệ C2 (trong (3.3.5) C2
Hãy để chúng tôi thực hiện một nghiên cứu chi tiết về mô hình này, sử dụng ký hiệu được chấp nhận rộng rãi nhất trong các tài liệu. Tân trang lại

Cơm. 3.2. Vị trí của các đường đẳng chính trong chân dung pha của động vật săn mồi thuộc hệ thống Volterra đối với các tỷ lệ thông số khác nhau: một- Về -
TỪ Tôi C2 C2
1, 3 - không ổn định, 2 - điểm kỳ dị ổn định; Trong -
1, 2, 3 - không ổn định, 4 - điểm kỳ dị ổn định có ý nghĩa
Hệ thống săn mồi-con mồi trong các ký hiệu này có dạng:

Chúng ta sẽ nghiên cứu các tính chất của các nghiệm đối với hệ (3.3.6) trên mặt phẳng pha N1
TRÊN2
Hệ thống có hai nghiệm tĩnh. Chúng dễ dàng xác định bằng cách cân bằng các cạnh bên phải của hệ thống bằng không. Chúng tôi nhận được: 
Do đó, các giải pháp tĩnh:

Chúng ta hãy xem xét kỹ hơn giải pháp thứ hai. Chúng ta hãy tìm tích phân đầu tiên của hệ (3.3.6) không chứa t. Nhân phương trình thứ nhất với -72, phương trình thứ hai với -71 và cộng các kết quả. Chúng tôi nhận được:
Bây giờ chúng ta chia phương trình đầu tiên cho N và nhân với € 2 và chia giây thứ hai cho JV 2 và nhân với e. Hãy thêm kết quả một lần nữa:
So sánh (3.3.7) và (3.3.8), chúng ta sẽ có:

Tích hợp, chúng tôi nhận được:

Đây là tích phân đầu tiên mong muốn. Do đó, hệ (3.3.6) là bảo toàn, vì nó có tích phân đầu tiên của chuyển động, một đại lượng là hàm của các biến của hệ N và N2 và không phụ thuộc vào thời gian. Tính chất này giúp ta có thể xây dựng một hệ thống các khái niệm cho hệ Volterra tương tự như cơ học thống kê (xem Chương 5), trong đó một vai trò thiết yếu được đóng bởi độ lớn của năng lượng của hệ, không thay đổi theo thời gian.
Đối với mọi cố định c> 0 (tương ứng với dữ liệu ban đầu nhất định), tích phân tương ứng với một quỹ đạo nhất định trên mặt phẳng N1 TRÊN2 , đóng vai trò là quỹ đạo của hệ thống (3.3.6).
Hãy xem xét một phương pháp đồ họa để xây dựng một quỹ đạo, do chính Volterra đề xuất. Lưu ý rằng vế phải của công thức (3.3.9) chỉ phụ thuộc vào D r 2 và vế trái chỉ phụ thuộc vào N. Chứng tỏ
Từ (3.3.9) nó theo sau rằng giữa X và Y có một mối quan hệ tỷ lệ thuận
Trên hình. 3.3 cho thấy các góc phần tư đầu tiên của bốn hệ tọa độ XOY, KHÔNG ĐƯỢC, N2 CON BÒ và D G 1 0N2 để chúng đều có một nguồn gốc chung.
Ở góc trên bên trái (góc phần tư KHÔNG)đồ thị của hàm (3.3.8) được xây dựng, ở phía dưới bên phải (góc phần tư N2 con bò)- đồ thị hàm số Y. Hàm đầu tiên có min tại Ni = và thứ hai - tối đa ở N2 = ?-
Cuối cùng, trong góc phần tư XOY xây dựng dòng (3.3.12) cho một số TỪ.
Đánh dấu một điểm N trên trục TRÊN. Điểm này tương ứng với một giá trị nhất định Y (N 1), dễ dàng tìm thấy bằng cách vẽ một đường vuông góc

Cơm. 3.3.
xuyên qua N cho đến khi nó giao với đường cong (3.3.10) (xem Hình 3.3). Lần lượt, giá trị của K (A ^) tương ứng với điểm M nào đó trên đoạn thẳng Y = cX và do đó một số giá trị X (N) = Y (N) / c có thể được tìm thấy bằng cách vẽ các đường vuông góc sáng và MD. Giá trị tìm được (điểm này được đánh dấu trong hình bằng chữ cái D) phù hợp với hai điểm R và G trên đường cong (3.3.11). Bằng những điểm này, vẽ đường vuông góc, chúng ta tìm thấy hai điểm cùng một lúc E " và E"nằm trên đường cong (3.3.9). Tọa độ của chúng là:

Vẽ vuông góc sáng, chúng tôi đã vượt qua đường cong (3.3.10) tại một điểm nữa TẠI.Điểm này tương ứng với R và Q trên đường cong (3.3.11) và giống nhau N và SCH.Điều phối Nđiểm này có thể được tìm thấy bằng cách thả vuông góc từ TẠI mỗi trục TRÊN. Vì vậy, chúng tôi nhận được điểm F " và F ”cũng nằm trên đường cong (3.3.9).
Đến từ một điểm khác N, theo cách tương tự, chúng ta thu được bốn điểm mới nằm trên đường cong (3.3.9). Ngoại lệ là dấu chấm Ni=? 2 / 72- Dựa vào đó, chúng ta chỉ nhận được hai điểm: Đến và L.Đây sẽ là điểm dưới và điểm trên của đường cong (3.3.9).
Không thể đến từ các giá trị N và từ các giá trị N2 . Đi từ N2 đến đường cong (3.3.11), sau đó tăng lên đường thẳng Y = cX, và từ đó băng qua đường cong (3.3.10), chúng ta cũng tìm thấy bốn điểm của đường cong (3.3.9). Ngoại lệ là dấu chấm Không =? 1 / 71- Dựa vào đó, chúng ta chỉ nhận được hai điểm: G và ĐẾN.Đây sẽ là điểm ngoài cùng bên trái và ngoài cùng bên phải của đường cong (3.3.9). Bằng cách hỏi khác nhau N và N2 và sau khi nhận đủ điểm, nối chúng với nhau, chúng ta sẽ dựng được đường cong (3.3.9).
Từ cấu trúc có thể thấy rằng đây là một đường cong khép kín chứa bên trong chính điểm 12 = (? 2/721? N yu và N20. Lấy một giá trị khác của C, tức là dữ liệu ban đầu khác, chúng ta nhận được một đường cong khép kín khác không giao với đường cong đầu tiên và cũng chứa điểm (? 2/721? 1/71) 1 bên trong chính nó. Do đó, họ quỹ đạo (3.3.9) là họ đường khép kín bao quanh điểm 12 (xem Hình 3.3). Chúng tôi khảo sát kiểu ổn định của điểm kỳ dị này bằng phương pháp Lyapunov.
Vì tất cả các thông số e 1,? 2, 71,72 là tích cực, dấu chấm (N [nằm trong góc phần tư dương của mặt phẳng pha. Sự tuyến tính hóa của hệ thống gần điểm này cho:

Nơi đây n (t) và 7i2 (N1, N2 :
Phương trình đặc trưng của hệ thống (3.3.13):

Các gốc của phương trình này hoàn toàn là tưởng tượng:

Vì vậy, nghiên cứu của hệ thống cho thấy rằng các quỹ đạo gần điểm kỳ dị được biểu diễn bằng các hình elip đồng tâm, và điểm kỳ dị chính nó là trung tâm (Hình 3.4). Mô hình Volterra đang được xem xét cũng có các quỹ đạo đóng cách xa điểm kỳ dị, mặc dù hình dạng của các quỹ đạo này đã khác với hình elip. Hành vi thay đổi Ni, N2 trong thời gian được hiển thị trong Hình. 3.5.

Cơm. 3.4.

Cơm. 3.5. Sự phụ thuộc của số lượng con mồi N tôi và động vật ăn thịt N2 từ lúc nào
Một điểm số ít của loại tâm là ổn định, nhưng không tiệm cận. Hãy sử dụng ví dụ này để chỉ ra nó là gì. Hãy để những rung động Ni (t) và LGgM xảy ra theo cách mà điểm đại diện di chuyển dọc theo mặt phẳng pha dọc theo quỹ đạo 1 (xem Hình 3.4). Tại thời điểm điểm ở vị trí M, một số lượng cá thể nhất định được thêm vào hệ thống từ bên ngoài N 2 sao cho điểm đại diện nhảy từ điểm Mđiểm A / ". Sau đó, nếu hệ lại tự động thì dao động Ni và N2 sẽ xảy ra với biên độ lớn hơn trước, và điểm đại diện di chuyển dọc theo quỹ đạo 2. Điều này có nghĩa là các dao động trong hệ không ổn định: chúng vĩnh viễn thay đổi các đặc tính của chúng dưới tác động bên ngoài. Theo đó, chúng ta xem xét các mô hình mô tả các chế độ dao động ổn định và chỉ ra rằng các chuyển động tuần hoàn ổn định tiệm cận được biểu diễn trên mặt phẳng pha bằng các chu kỳ giới hạn.
Trên hình. 3.6 cho thấy các đường cong thực nghiệm - sự dao động về số lượng động vật mang lông ở Canada (theo Hudson's Bay Company). Các đường cong này được xây dựng trên cơ sở dữ liệu về số lượng da thu hoạch được. Các thời kỳ dao động về số lượng của thỏ rừng (con mồi) và linh miêu (động vật ăn thịt) là xấp xỉ nhau và có thứ tự từ 9-10 năm. Đồng thời, theo quy luật, số lượng thỏ rừng tối đa đi trước số lượng linh miêu tối đa một năm.
Hình dạng của những đường cong thực nghiệm này kém chính xác hơn nhiều so với những đường cong lý thuyết. Tuy nhiên, trong trường hợp này, đủ để mô hình đảm bảo sự trùng hợp của các đặc điểm quan trọng nhất của đường cong lý thuyết và thực nghiệm, tức là giá trị biên độ và độ lệch pha giữa các dao động của số lượng vật ăn thịt và con mồi. Một thiếu sót nghiêm trọng hơn nhiều của mô hình Volterra là tính không ổn định của các nghiệm đối với hệ phương trình. Thật vậy, như đã đề cập ở trên, bất kỳ sự thay đổi ngẫu nhiên nào về sự phong phú của một loài hoặc loài khác, theo mô hình, sẽ dẫn đến sự thay đổi biên độ dao động của cả hai loài. Đương nhiên, trong điều kiện tự nhiên, động vật phải chịu vô số tác động ngẫu nhiên như vậy. Qua các đường cong thí nghiệm có thể thấy, biên độ dao động số lượng loài ít thay đổi theo từng năm.
Mô hình Volterra là một mô hình tham chiếu (cơ bản) cho sinh thái toán học ở mức độ tương tự như mô hình dao động điều hòa là cơ bản cho cơ học cổ điển và lượng tử. Với sự trợ giúp của mô hình này, dựa trên những ý tưởng rất đơn giản về bản chất của các mẫu mô tả hành vi của hệ thống, thuần túy là toán học
Chương 3

Cơm. 3.6. Đường cong động học của sự phong phú của động vật mang lông Theo Công ty Hudson's Bay Fur (Seton-Thomson, 1987), một kết luận được rút ra bằng phép tính về bản chất định tính của hành vi của một hệ thống như vậy - về sự hiện diện của sự biến động dân số trong một hệ thống như vậy. Nếu không có việc xây dựng một mô hình toán học và sử dụng nó, thì một kết luận như vậy sẽ là không thể.
Ở dạng đơn giản nhất mà chúng ta đã xem xét ở trên, hệ thống Volterra có hai khuyết điểm cơ bản và có liên quan lẫn nhau. "Sự loại bỏ" của họ được dành cho các tài liệu sinh thái học và toán học mở rộng. Thứ nhất, việc đưa vào mô hình của bất kỳ yếu tố bổ sung nào, nhỏ tùy ý, thay đổi về mặt chất lượng hành vi của hệ thống. Hạn chế “sinh học” thứ hai của mô hình là nó không bao gồm các đặc tính cơ bản vốn có trong bất kỳ cặp quần thể nào tương tác theo nguyên tắc kẻ săn mồi - con mồi: ảnh hưởng của sự bão hòa của kẻ thù, nguồn tài nguyên hạn chế của kẻ săn mồi và con mồi thậm chí. với lượng con mồi dư thừa, khả năng có thể có số lượng con mồi tối thiểu cho kẻ săn mồi, v.v.
Để loại bỏ những nhược điểm này, các sửa đổi khác nhau của hệ thống Volterra đã được các tác giả khác nhau đề xuất. Điều thú vị nhất trong số chúng sẽ được xem xét trong phần 3.5. Ở đây chúng tôi chỉ tập trung vào một mô hình có tính đến những hạn chế của bản thân trong sự tăng trưởng của cả hai quần thể. Ví dụ của mô hình này cho thấy rõ bản chất của các giải pháp có thể thay đổi như thế nào khi các thông số hệ thống thay đổi.
Vì vậy, chúng tôi coi hệ thống

Hệ thống (3.3.15) khác với hệ thống đã xét trước đây (3.3.6) bởi sự hiện diện của các số hạng có dạng -7 ở vế phải của phương trình uNf,
Các thành viên này phản ánh một thực tế là quần thể con mồi không thể phát triển vô hạn ngay cả khi không có động vật ăn thịt do nguồn thức ăn hạn chế, phạm vi tồn tại hạn chế. Những "giới hạn bản thân" tương tự cũng được áp đặt cho quần thể động vật ăn thịt.
Để tìm số lượng cố định của các loài iVi và N2 tương đương với không các phần bên phải của phương trình của hệ (3.3.15). Các giải pháp với số lượng động vật ăn thịt hoặc con mồi bằng 0 sẽ không khiến chúng ta quan tâm. Do đó, hãy xem xét một hệ thức đại số
phương trình  Quyết định của cô ấy
Quyết định của cô ấy
cho chúng ta tọa độ của điểm kỳ dị. Ở đây, điều kiện tích cực của các số đứng yên nên được đặt trên các tham số của hệ thống: N> 0 và N2 > 0. Các gốc của phương trình đặc trưng của một hệ thống được tuyến tính hóa trong vùng lân cận của một điểm kỳ dị (3.3.16):

Có thể thấy từ biểu thức cho các số đặc trưng rằng nếu điều kiện
khi đó số lượng vật ăn thịt và con mồi thực hiện dao động tắt dần trong thời gian, hệ có điểm kỳ dị khác 0 và trọng tâm ổn định. Chân dung pha của một hệ thống như vậy được thể hiện trong Hình. 3,7 a.
Giả sử rằng các tham số trong bất đẳng thức (3.3.17) thay đổi giá trị của chúng theo cách mà điều kiện (3.3.17) trở thành một đẳng thức. Khi đó các số đặc trưng của hệ thống (3.3.15) bằng nhau, và điểm kỳ dị của nó sẽ nằm trên ranh giới giữa các vùng của các điểm và nút ổn định. Khi dấu của bất đẳng thức (3.3.17) được đảo ngược, điểm kỳ dị trở thành một nút ổn định. Chân dung pha của hệ thống cho trường hợp này được thể hiện trong Hình. 3,76.
Như trong trường hợp của một tập hợp đơn lẻ, một mô hình ngẫu nhiên có thể được phát triển cho mô hình (3.3.6), nhưng nó không thể được giải quyết một cách rõ ràng. Do đó, chúng ta tự giới hạn mình trong những cân nhắc chung chung. Ví dụ, giả sử rằng điểm cân bằng nằm ở một khoảng cách nào đó từ mỗi trục. Sau đó, đối với quỹ đạo pha mà các giá trị của JVj, N2 vẫn đủ lớn, một mô hình xác định sẽ khá thỏa đáng. Nhưng nếu một lúc nào đó


Cơm. 3.7. Chân dung pha của hệ thống (3.3.15): một - khi mối quan hệ (3.3.17) giữa các tham số được thỏa mãn; b- khi thực hiện mối quan hệ nghịch đảo giữa các tham số
quỹ đạo pha, bất kỳ biến số nào không lớn lắm, khi đó dao động ngẫu nhiên có thể trở nên đáng kể. Chúng dẫn đến thực tế là điểm đại diện sẽ di chuyển đến một trong các trục, đồng nghĩa với sự tuyệt chủng của các loài tương ứng. Do đó, mô hình ngẫu nhiên hóa ra không ổn định, vì sự "trôi dạt" ngẫu nhiên sớm hay muộn dẫn đến sự tuyệt chủng của một trong các loài. Trong loại mô hình này, động vật ăn thịt cuối cùng chết đi, do ngẫu nhiên hoặc do quần thể con mồi của nó bị loại bỏ trước. Mô hình ngẫu nhiên của hệ thống động vật ăn thịt-con mồi giải thích tốt các thí nghiệm của Gause (Gause, 1934; 2000), trong đó ciliates Paramettum candatum phục vụ như một con mồi cho một con ciliate khác Didinium nasatum- động vật ăn thịt. Số lượng cân bằng dự kiến theo phương trình xác định (3.3.6) trong các thí nghiệm này chỉ xấp xỉ năm cá thể của mỗi loài, vì vậy không có gì đáng ngạc nhiên trong thực tế là trong mỗi thí nghiệm lặp lại, động vật ăn thịt hoặc con mồi chết khá nhanh (và sau đó động vật ăn thịt).).
Vì vậy, việc phân tích các mô hình Volterra về sự tương tác giữa các loài cho thấy rằng, mặc dù có rất nhiều kiểu hành vi của các hệ thống như vậy, nhưng không thể có sự biến động quần thể nào chưa được đánh dấu trong mô hình của các loài cạnh tranh. Trong mô hình kẻ săn mồi - con mồi, các dao động không dấu xuất hiện do sự lựa chọn dạng đặc biệt của phương trình mô hình (3.3.6). Trong trường hợp này, mô hình trở nên không thô, điều này cho thấy sự vắng mặt của các cơ chế trong một hệ thống tìm cách duy trì trạng thái của nó. Tuy nhiên, những biến động như vậy được quan sát trong tự nhiên và thực nghiệm. Nhu cầu giải thích lý thuyết của họ là một trong những lý do để xây dựng mô tả mô hình ở dạng tổng quát hơn. Phần 3.5 được dành để xem xét các mô hình tổng quát như vậy.